

イソギンチャクの体の構造に“相称性の二刀流”を発見
左右相称と放射相称の両方をこなす体づくりの数理モデル
概要
理化学研究所(理研)生命機能科学研究センター形態進化研究チームのサフィエ・エスラ・サルペル訪問研究員(日本学術振興会特別研究員-PD、大阪大学大学院招へい研究員)、平井珠美テクニカルスタッフ、倉谷滋チームリーダー(開拓研究本部倉谷形態進化研究室主任研究員)、大阪大学大学院理学研究科の藤本仰一准教授らの共同研究グループ※は、「タテジマイソギンチャクに左右相称の個体と放射相称の個体が混在することを発見し、両者の体づくりを共通の仕組みで説明する数理モデルを提案しました。
本研究により、現生のほとんどの動物に共通する左右相称性と、進化において祖先的な動物である刺胞動物門(クラゲ、イソギンチャクなど)で見られる放射相称性の体づくりの仕組みの関係や、相称性がどのように進化してきたかについての理解が深まることが期待できます。
これまで、左右相称性と放射相称性が同じ生物で現れることはないとされてきました。しかし今回、共同研究グループは、タテジマイソギンチャクが左右相称と放射相称の2タイプの個体を持つ、いわば“相称性の二刀流”をこなす珍しい種であることを発見しました。体内構造を解析した結果、イソギンチャク特有の器官である「管溝」が、左右相称個体では一つ、放射相称個体では二つあることが分かりました。さらに数理モデルを用いたシミュレーションから、個体の相称性は再生の初期段階における管溝胃嚢 の有無により決まることが示されました。
本研究は、科学雑誌『Zoological Letters』オンライン版(9月6日付)に掲載されました。
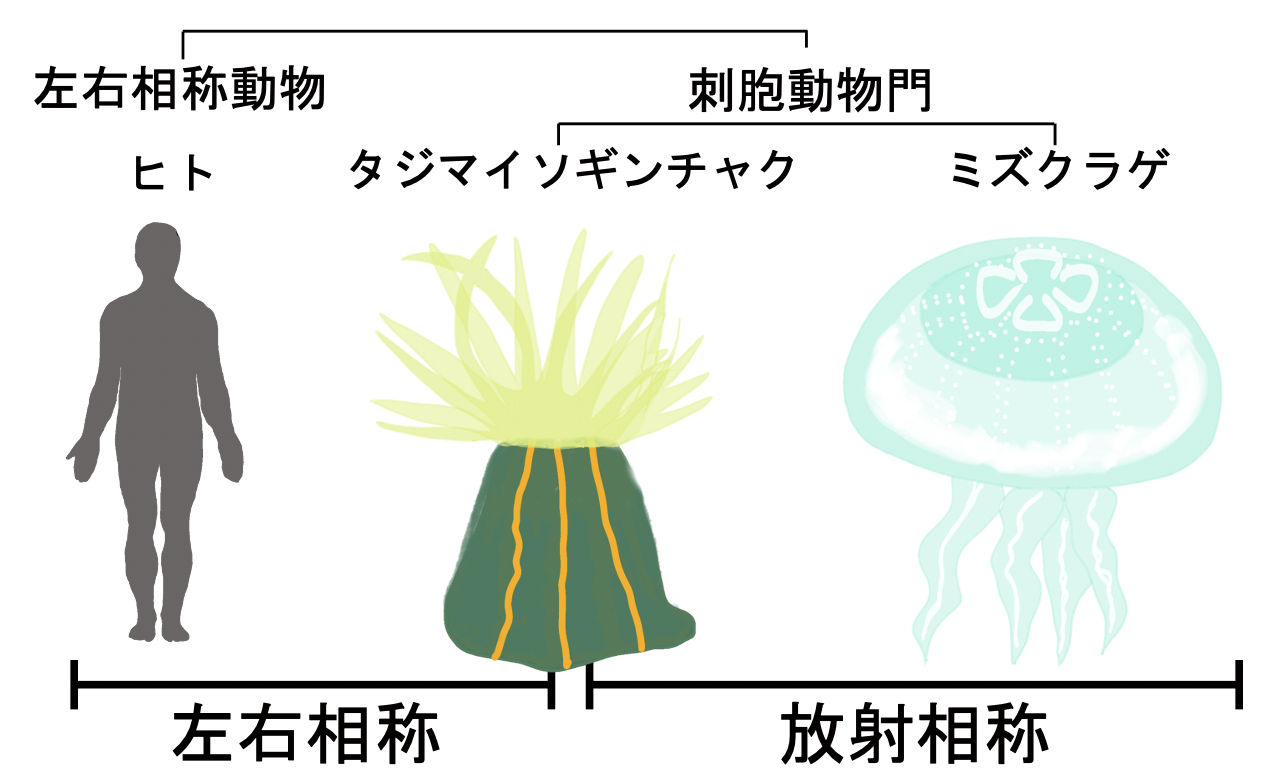
図 左右相称と放射相称の“相称性の二刀流”をこなすタテジマイソギンチャク
※共同研究グループ
理化学研究所 生命機能科学研究センター
形態進化研究チーム
チームリーダー 倉谷 滋 (くらたに しげる)
(開拓研究本部 倉谷形態進化研究室 主任研究員)
訪問研究員 サフィエ・エスラ・サルペル(Safiye Esra Sarper)
(日本学術振興会特別研究員-PD、大阪大学大学院招へい研究員)
テクニカルスタッフ 平井 珠美 (ひらい たまみ)
網膜再生医療研究開発プロジェクト
客員研究員 松山 オジョス 武(Matsuyama Hoyos Takesi)
大阪大学 大学院理学研究科 生物科学専攻 理論生物学研究室
准教授 藤本 仰一 (ふじもと こういち)
背景
クラゲは宇宙人など未知の生物のモチーフとして数々の小説や映画で使われてきました。これはクラゲを含む刺胞動物門の動物の体の構造が、ヒトや身近な動物と大きく異なることが原因だと考えられます。このような動物の体の構造の違いがどのように生み出されるかは、いまだに解明されていません。
ヒトを含む多くの動物の体の構造は、体の中心線を通る面(正中面)を挟んで左右対称になるように配置されています。これは「左右相称性」と呼ばれ、ほとんどの動物で見られる代表的な特徴です。一方、体を対称に分ける面が複数存在する「放射相称性」は、進化において祖先的な動物の特徴であると考えられています。現生の動物では、クラゲ、ヒドラ、イソギンチャクなどを含む刺胞動物門に属する種の多くが放射相称の構造を持っています(図1)。
動物の進化において、体の構造の相称性が変わることは大きな転換であり、これまで多くの研究がなされてきました。刺胞動物門の中でもイソギンチャクの仲間には、左右相称性を示す種と放射相称性を示す種が存在しており、相称性の進化を研究する対象として適しています。特に最近、脊椎動物の発生過程で、正中面に沿った体の構造を決定する遺伝子群が、左右相称性を示すイソギンチャクの発生過程でも似た発現パターンを持つことが明らかになり注)、刺胞動物門とそれ以外の動物に左右相称な体づくりの共通性があると考えられるようになりました。
しかし、放射相称性を示すイソギンチャクの体づくりの仕組みに関する解析は進んでおらず、放射相称性と左右相称性の違いがどのように生み出されるかもよく分かっていません。そこで共同研究グループは、放射相称性と左右相称性それぞれの形成過程を比較するため、これに適したイソギンチャクの探索を試みました。
注)He, S. et al. An axial Hox code controls tissue segmentation and body patterning in Nematostella vectensis. Science 361, 1377–1380 (2018).
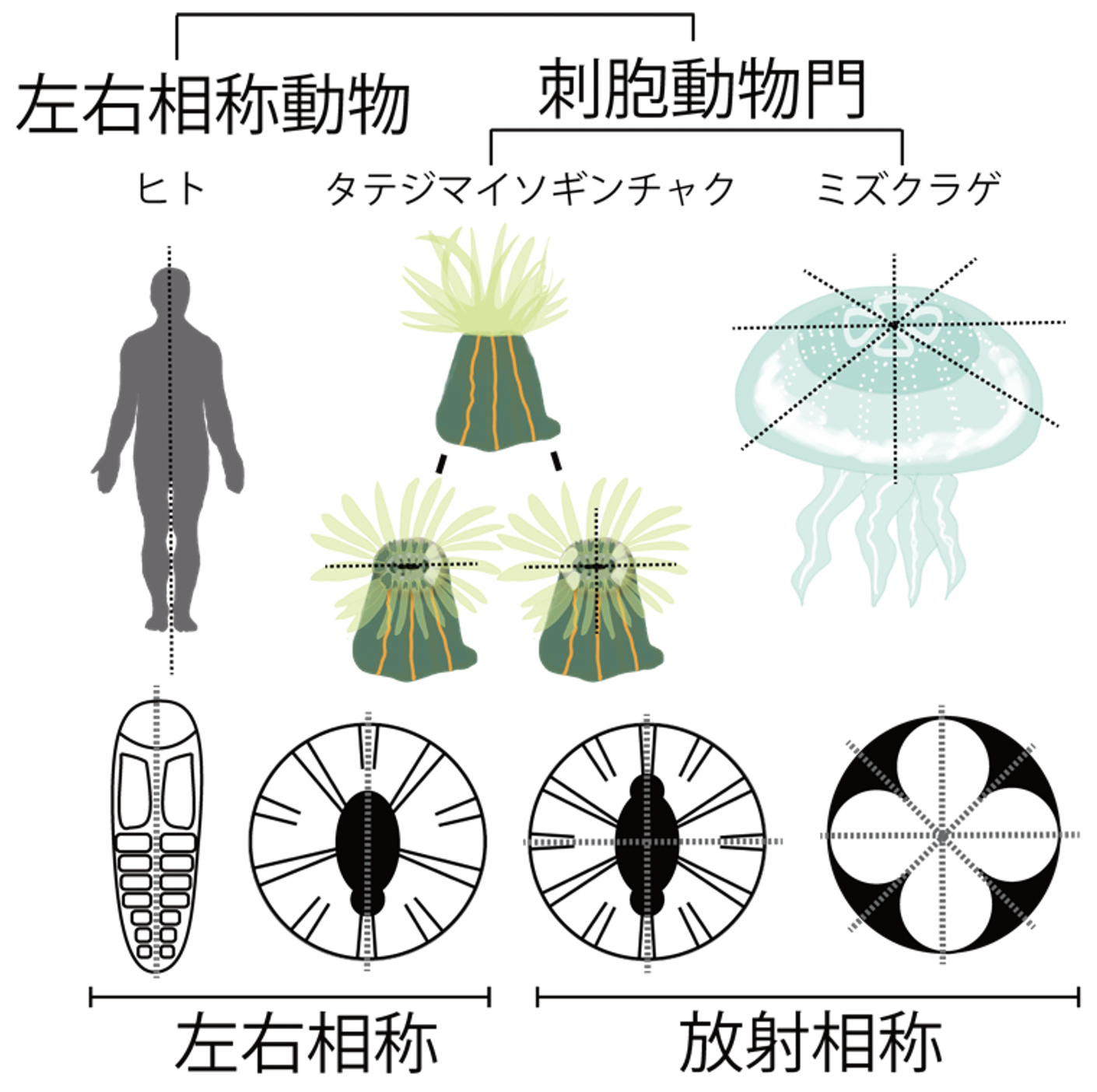
図1. 動物界で見られる相称性のタイプ
脊椎動物や節足動物など、体の基本構造が左右相称である動物は左右相称動物と総称される。一方、クラゲ、イソギンチャク、サンゴなどが属する刺胞動物門では、多くの種が放射相称であるが、一部に左右相称の種を含む。なかでも本研究で見いだしたタテジマイソギンチャクは、種内にいずれかの相称性を持つ個体が混在する。点線は対称軸を指す。
研究手法と成果
共同研究グループは、左右相称性を示す種と放射相称性を示す種の両方が報告されているイソギンチャクの仲間に注目した結果、「タテジマイソギンチャク」が種内変異(個体差)として両方の相称性が混在している種であることを発見しました。
タテジマイソギンチャクは日本に広く分布しており、名前の由来である縦に伸びるオレンジ色の縞模様が特徴です(図2a)。潮間帯の岩の隙間などに見られ、口の反対側(反口側)を岩に付着させて生活しています。潮が満ちてくると、波で運ばれてくるプランクトンや小エビを口側に生えている触手で捕まえ、口腔に入れて食べます。口腔の側面には、体内に入る海水の量を調節する「管溝」器官が存在しています(図2b, c)。体内には胃の働きをする「胃嚢」が複数存在し、口腔の周りにピザを切り分けたような放射状に配置されています(図2c, d)。
本研究ではこれらの胃嚢を、大きさや管溝に対する位置によって主な3種類に分類しました(第1胃嚢、第2胃嚢、管溝胃嚢)(図2 c, d)。この3種類の胃嚢の配置には個体差があり、これによって個体の相称性が決まります。
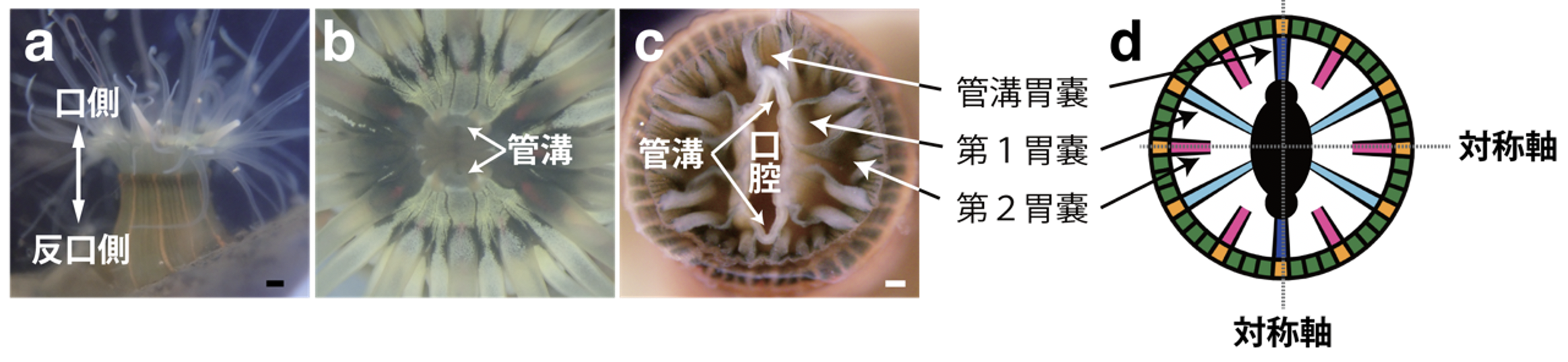
図2. タテジマイソギンチャクの体の構造
a) タテジマイソギンチャクの外観。反口側(写真下側)で岩に付着し、口側の触手で食べ物を取る。オレンジ色の縦の縞が明瞭に見える。スケールバーは2mm。
b) 口側から見た口腔。管溝の周りには、扇形に広がる薄緑色の構造が見られる。
c) 口腔の水平断面図。スケールバーは200マイクロメートル(μm、1μmは1,000分の1mm)。
d) 胃嚢の配置パターンを示した模式図。管溝に接続する胃嚢(管溝胃嚢:濃い青)、長さの異なる第1胃嚢(青)、第2胃嚢(ピンク)の配置により、放射相称か左右相称かを判断できる。この模式図は、管溝胃嚢を結ぶ縦の軸と、それに直行する第2胃嚢を結ぶ横の軸の二つの対称軸がある二放射相称を示す。
次に、これらの体内構造と、外部から観察可能な形態的特徴の対応関係を調べたところ、オレンジ色の縞は第1胃嚢、第2胃嚢と管溝胃嚢の場所を指していることが分かりました。さらに、口腔の周りに存在する扇形の薄緑色素パターンは、管溝の位置を要として広がることを発見しました。
野外から採取した114個体について、縞の数や色素パターンを解析して3種類の胃嚢の配置や数を調べたところ、放射相称(二放射相称)個体が最も多く、その次には左右相称個体が多く見つかりました。さらに、左右相称個体は必ず一つの管溝を持ち、放射相称個体は必ず二つの管溝を持つことが明らかになりました(図3)。これらの結果から、管溝およびそれに接続する管溝胃嚢が、体の対称構造を作る仕組みに重要な役割を担っていることが示されました。
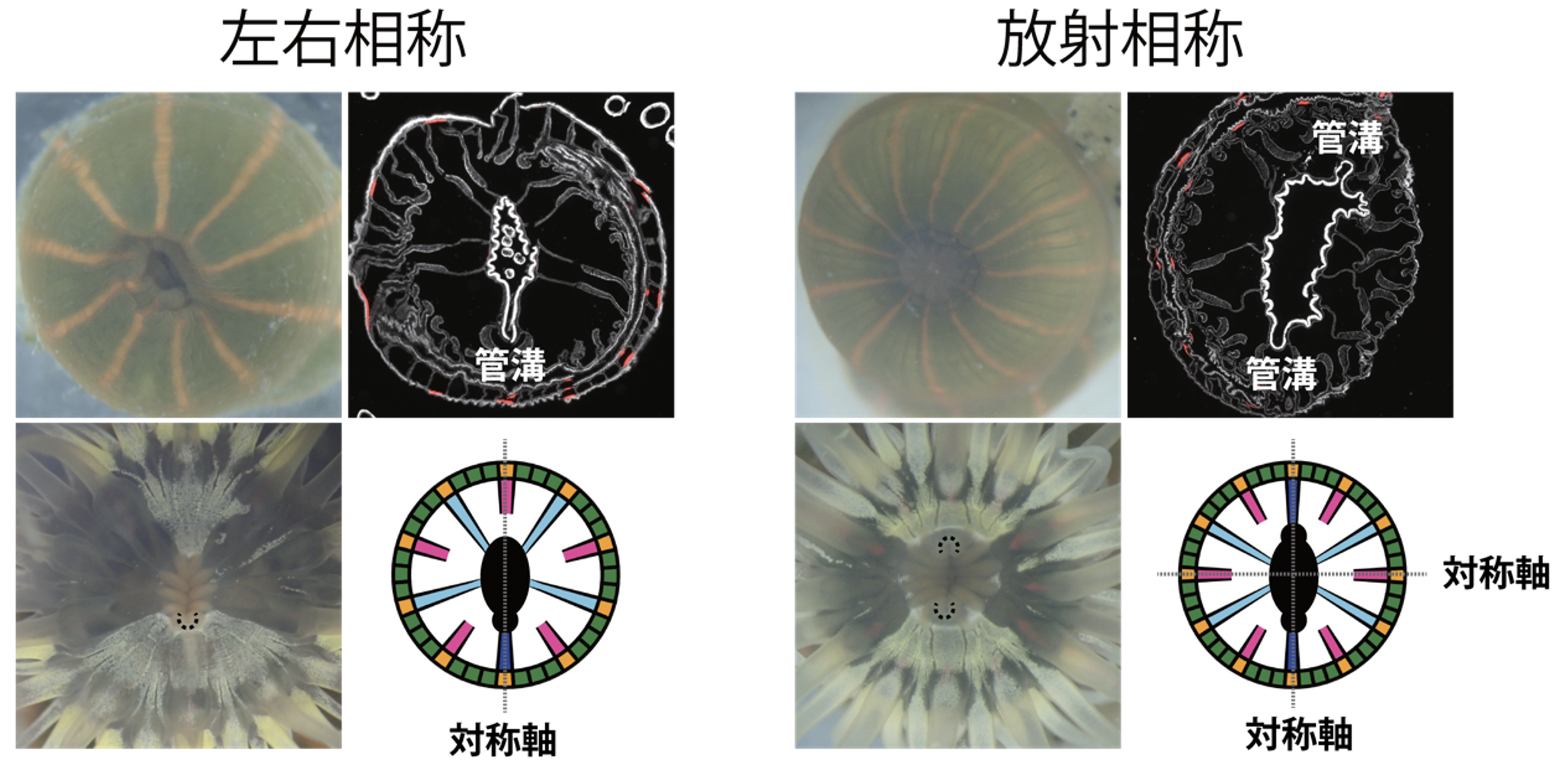
図3. タテジマイソギンチャク個体の“相称性の二刀流”
左右相称個体(左)と放射相称個体(右)。それぞれ、左上から時計回りに、外観、断面像、体内構造の模式図、口腔の拡大像を示した。左下写真内の黒点の円弧は管溝を指す。
タテジマイソギンチャクはモデル動物ではないため、遺伝子解析が困難です。そこで、今回は数理モデルを用いたコンピュータシミュレーションを行い、管溝を中心とする体づくりの仕組みを予測しました。タテジマイソギンチャクは有性生殖に加えて、反口側の体の一部が分裂し、その断片から新しい個体が再生する無性生殖も行います。この無性生殖による再生過程では、管溝胃嚢の位置が最初に決まり、その後、第1胃嚢と第2胃嚢が互い違いにほぼ規則的な間隔で出現することが知られています。
この知見を基に、第1胃嚢と第2胃嚢はそれぞれ自分自身の形成と相手の形成を抑制する因子を分泌し、管溝胃嚢は第1胃嚢と第2胃嚢の形成を促進・抑制する因子を分泌するという側方促進・抑制モデルを構築しました(図4)。シミュレーションの結果、同じ分子メカニズムから放射相称と左右相称の両方の相称性の形成が可能であることが示されました。また、分裂した断片に含まれる胃嚢の種類に応じて再生個体の相称性が異なり、管溝胃嚢を含む断片からは放射相称個体が、第1胃嚢もしくは第2胃嚢を含む断片からは左右相称個体が再生されることも分かりました。
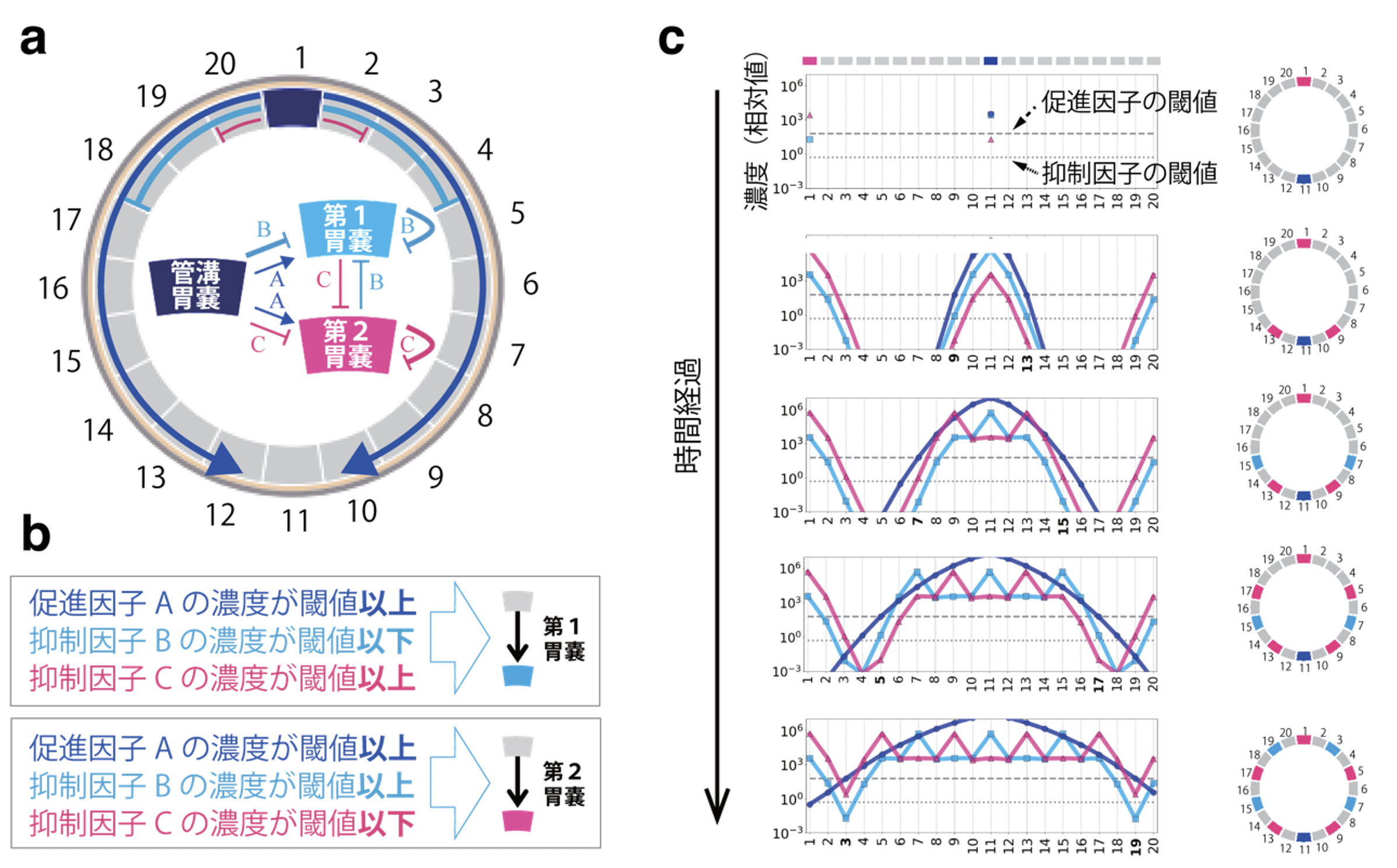
図4. タテジマイソギンチャクの再生過程における胃嚢の領域化と数理モデル
a) 再生により再構築された体内構造の模式図。最初の断片に含まれていた胃嚢の位置を1とし、時計回りに番号をふった。このとき、最初の断片の反対側の胃嚢(11)には必ず管溝胃嚢が形成されるが、他の領域が何になるかは未決定とした。さらにこれらの領域は再生の過程で、それぞれの胃嚢から分泌される拡散性の促進因子(A)と抑制因子(B、C)の作用を受けて決定される、側方促進・抑制モデルを構築した(模式図中にこれらの関係を矢印で示す)。
b) 側方促進・抑制モデルでは、 3種類の因子(A、B、C)の作用を受けた未決定の領域は、それぞれの濃度に依存して第1胃嚢になるか、第2胃嚢になるかが決定される。
c) 最初の断片に第2胃嚢が含まれていた場合の再生シミュレーション。左のグラフは時間経過とともに変化するA、B、Cの濃度勾配を示す。右の模式図は、各領域のA、B、Cの濃度の関係で第1胃嚢と第2胃嚢が決定されていく様子を示す。
以上の結果から、タテジマイソギンチャクで放射相称と左右相称の2タイプの個体が存在するのは、再生の初期段階における管溝胃嚢の有無によるとの仮説が立てられます(図5)。
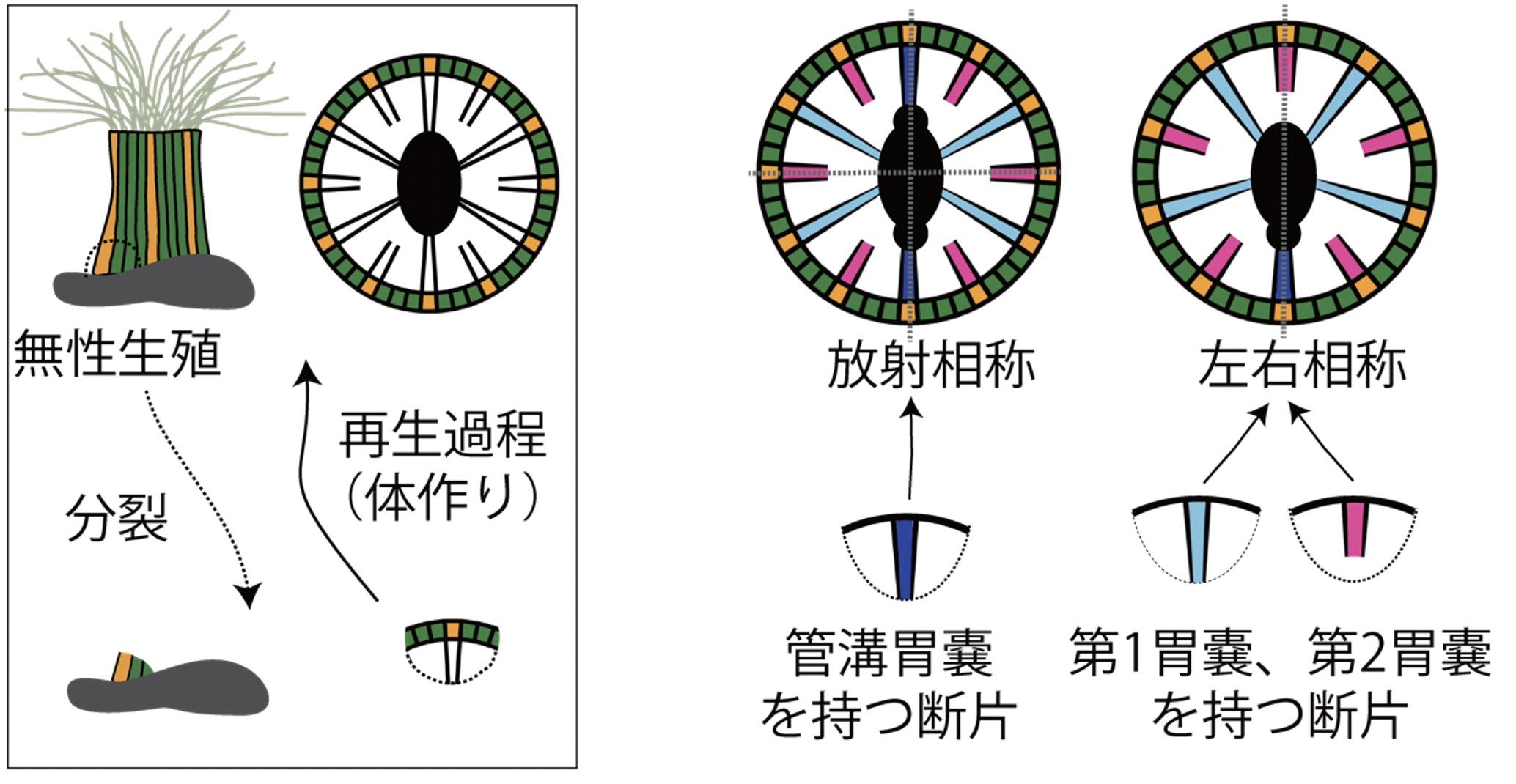
図5. タテジマイソギンチャクの無性生殖で生じる二つの相称性
採取された個体に多く見られる左右相称と放射相称の体の構造。シミュレーションの結果から、これらの違いは、無性生殖の際に管溝胃嚢を持つ断片から再生したか、それ以外の胃嚢を持つ断片から再生したかによって決まることが示唆された。
今後の期待
動物の相称性の進化や左右相称動物の起源については、多くの謎が残されています。本研究では、左右相称個体と放射相称個体が共存している珍しい種、タテジマイソギンチャクに着目し、これら二つの異なる相称性が同じ遺伝子のプログラムからでも出現し得る仕組みを数理モデルから提案しました。
今後、タテジマイソギンチャクの再生過程をさらに詳しく解析することで、この予測の検証が期待されます。さらに、無性生殖後の再生や有性生殖後の発生過程を他の種を含めて広く調べることで、相称性の進化に関する新たな知見が得られると期待できます。
特記事項
論文情報
<タイトル>
Polymorphism in the symmetries of gastric pouch arrangements in the sea anemone Diadumene lineata
<著者名>
Safiye E. Sarper, Tamami Hirai, Take Matsuyama, Shigeru Kuratani, and Koichi Fujimoto
<雑誌>
Zoological letters
<DOI>
10.1186/s40851-021-00180-0
本研究は、日本学術振興会(JSPS)科学研究費補助金新学術領域研究(研究領域提案型)「脊椎動物の筋骨格系の形態進化に見る制約と方向性(領域代表者:倉谷滋)」、同「進化のゆらぎ応答理論の確立と多階層・発生過程への展開(研究代表者:金子邦彦、研究分担者:藤本仰一)」による支援を受けて行われました。
用語説明
- タテジマイソギンチャク
学名Diadumene lineata。日本で普通に見られる小型のイソギンチャク 。有性生殖のほか、岩に付着する足盤が分裂して増える無性生殖も行う。
- 左右相称、放射相称
動物の体の体制について、前と後、背と腹が区別され、外見的に左側と右側の形が等しいものを左右相称と呼ぶ。一方、一つの対称軸を含む相称面が複数存在するものを放射相称と呼ぶ。
- 刺胞動物門
動物分類の最も大きな単位「門」のうち、クラゲ、ヒドラ、サンゴ、イソギンチャクなどを含むグループ。
- 管溝、胃嚢
イソギンチャク特有の器官の名称。イソギンチャクの口は細長く伸びており、その端に存在する溝状の構造を管溝と呼ぶ。管溝には繊毛を持つ細胞が並んでおり、体内に入る海水の量を調節する。胃嚢は、隔膜によって仕切られた胃の空間。
- 潮間帯
海の生態環境のうち、潮の満ち引きにより海面からの露出と水没を繰り返す場所を指す。
- 側方促進・抑制モデル
胃嚢の規則的な配置を説明するために本研究で立てられたモデル。動物の発生過程では、ある細胞が分泌した因子の拡散により、他の細胞の分化が促進あるいは抑制される例がよく知られており、これをタテジマイソギンチャクの再生過程の仕組みのモデルとして応用した。