
根が適切な間隔で分岐する仕組みに働く植物ペプチドを発見
ポイント
・植物の根を分岐させる「側根」を作り出す細胞(側根創始細胞 )が、すぐ近くに生じないように働くペプチドTOLS2(トルス2)を、モデル植物シロイヌナズナから発見した。
・TOLS2ペプチドは側根創始細胞で産生され、植物ホルモンのオーキシン とLBD16タンパク質(転写因子)によってその発現が誘導される。
・TOLS2ペプチドは、側根創始細胞の周囲の細胞にあるRLK7受容体タンパク質に受容されると、側根創始細胞の形成を抑制する遺伝子を活性化する。
・本研究で発見したTOLS2ペプチド-RLK7受容体を介した経路は、「側根形成の側方抑制」の仕組みに働くことが初めて示されたペプチドホルモン 応答経路である。
概要
神戸大学大学院理学研究科・深城英弘教授、豊倉浩一研究員(現・大阪大学・日本学術振興会特別研究員)、郷達明特命助教(現・奈良先端科学技術大学院大学・助教)らと、名古屋大学大学院理学研究科・松林嘉克教授、篠原秀文助教、および奈良先端科学技術大学院大学、大阪大学大学院理学研究科・藤本仰一准教授、東京大学大学院理学系研究科・近藤侑貴助教の共同研究グループは、モデル植物シロイヌナズナ を用いて、根を分岐させる細胞(側根創始細胞)が適切な間隔で生じる仕組みに働くペプチドとその受容体を明らかにしました。
今後、根を分岐させる仕組みが解明されれば、将来さまざまな農作物や樹木において、土壌における根の張り方を人為的に制御することが可能になることが期待されます。
この研究成果は、2018年12月21日(米国12月20日)に、国際学術雑誌「Developmental Cell」オンライン版に掲載されました。
研究の背景
植物は、土の中で新しい根を次々と分岐させることによって、生育環境に適した形の根系(植物体の地下部全体)を発達させます。根系は、主にすでに作られた根の内部組織から作られる側根によって形づくられます。側根は発芽後に多数作られ、根系の大部分を占めるため、側根は根系の形状パターンに大きな影響を及ぼします。
側根は、根の分裂組織から少し離れた部位で、規則的な間隔をもって生じる「側根創始細胞」とよばれる数個の細胞から作られます。これまでモデル植物のシロイヌナズナを用いた研究から、側根創始細胞は、根端の分裂組織から少し離れた領域に規則的に生じるオーキシン応答が高い部位(プレブランチサイト:分岐予定部位)から形成されることがわかっていました。しかし、このプレブランチサイトの細胞群から、どのような仕組みで数個の側根創始細胞が生じるのかその仕組みは不明でした。
一方、本研究グループによって、オーキシンによって側根創始細胞で誘導される転写因子LBD16には、側根創始細胞の周囲の細胞が側根創始細胞になるのを抑制する働きがあることが示唆されていました。しかし、その仕組みについてはわかっていませんでした。
研究の内容
本研究グループでは、モデル植物のシロイヌナズナにおいて、側根形成開始を制御する転写因子LBD16によって活性化する下流遺伝子を探索しました。その結果、低分子分泌型ペプチドをコードするTARGET OF LBD SIXTEEN2 (TOLS2)/PAMP-INDUCED SECRETED PEPTIDE-LIKE3遺伝子(以下、TOLS2遺伝子)を同定することに成功しました。
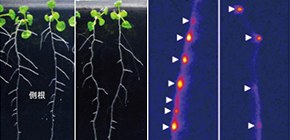
また、TOLS2遺伝子は側根創始細胞や側根原基で主に発現しますが、TOLS2を過剰に発現するシロイヌナズナでは、DR5 とよばれるマーカー遺伝子を発現する側根創始細胞の数が減少し、形成される側根数も減少しました (図1) 。
これらの結果から、TOLS2遺伝子には側根創始細胞の形成を抑制する働きがあることが示唆されました。TOLS2遺伝子は86アミノ酸からなるポリペプチドをコードしますが、そのアミノ酸配列から、十数アミノ酸からなるペプチドホルモンをコードすることが予想されました。そこで、TOLS2過剰発現植物から培養液中に分泌される物質をナノ液体クロマトグラフィー/質量分析計(Nano-LC-MS)で解析を行い、成熟型TOLS2ペプチドの構造が、水酸化プロリンを含む11アミノ酸からなることを明らかにしました。実際、この成熟型TOLS2ペプチドを人工合成して野生型シロイヌナズナに添加すると、側根創始細胞の数が減少し、形成される側根数も減少することが確認されました (図2) 。これらの結果から、成熟型TOLS2ペプチドには側根創始細胞の形成を抑制する働きがあることが明らかとなりました。
一般に、植物のペプチドホルモンはロイシンリッチリピート型受容体キナーゼ(LRR-RK)群とよばれるタンパク質群のいずれかによって受容されることが知られています。そこで、このグループに属する受容体キナーゼ遺伝子が欠損した複数のシロイヌナズナ変異体を調べたところ、RECEPTOR-LIKE7(RLK7)受容体に欠損のあるrlk7変異体において、TOLS2ペプチドによる側根形成の抑制が全く起こらないことを見出しました。また、生化学的な結合実験によって、TOLS2ペプチドがRLK7タンパク質の細胞外受容ドメイン(ロイシンリッチリピート領域)と結合することが示されました。これらの結果から、TOLS2ペプチドの受容体がRLK7であることが明らかとなりました。根においてRLK7タンパク質は内鞘(側根創始細胞が生じる細胞層)、内皮、皮層で発現しますが、側根創始細胞では発現がみられませんでした。おそらく側根創始細胞の周辺の細胞でのみ、側根形成を抑制していると考えられます。
次に、CRISPR/Cas9法 を用いたゲノム編集技術によって作出したTOLS2遺伝子とTOLS2ペプチドに類似したPIP2ペプチドをコードする遺伝子の両方に欠損のあるpip2 tols2二重変異体、およびTOLS2ペプチドを受容できないrlk7変異体では、側根創始細胞がどのように形成されるのかを調べました。その結果、これらの変異体では側根創始細胞の数が多く、また野生型よりも短い間隔で生じていました。さらに、経時的に観察すると野生型と変異体のどちらの根においても、時折、側根創始細胞のマーカーであるDR5活性のある部位が二つ近接して生じる場合が観察されました。しかし、野生型では片方だけがDR5活性を強め、もう片方の活性を弱めるケースが多いのに対し、rlk7変異体では両方のDR5活性が強い状態を維持する傾向があることが明らかになりました。これらの結果からも、TOLS2ペプチドとRLK7受容体が、側根創始細胞を生じる間隔を適切に保つ仕組みに必要であることが示されました。
このような一連の解析から本研究グループは、シロイヌナズナはオーキシンに応答してTOLS2ペプチドを側根創始細胞で誘導することで、RLK7受容体を介して近傍における側根創始細胞形成を非細胞自律的な方法で抑制する(側根形成の側方抑制)、という仕組みを提唱しました (図3) 。

図1 TOLS2遺伝子の過剰発現は側根創始細胞および側根の数を減少させる。
A:シロイヌナズナ野生型(Col)(左)とTOLS2過剰発現体(右)の10日目芽生え。スケールは1cm。
B:シロイヌナズナ野生型(Col)(左)とTOLS2過剰発現体(右)の根におけるDR5:LUC遺伝子の発現。白矢じりは側根創始細胞を示す。スケールは1cm。

図2 TOLS2ペプチドは側根の数を減少させる。
コントロールおよびTOLS2ペプチド(100nMおよび1M濃度)を含む培地で生育させたシロイヌナズナ野生型(Col)の10日目芽生え。スケールは1cm。
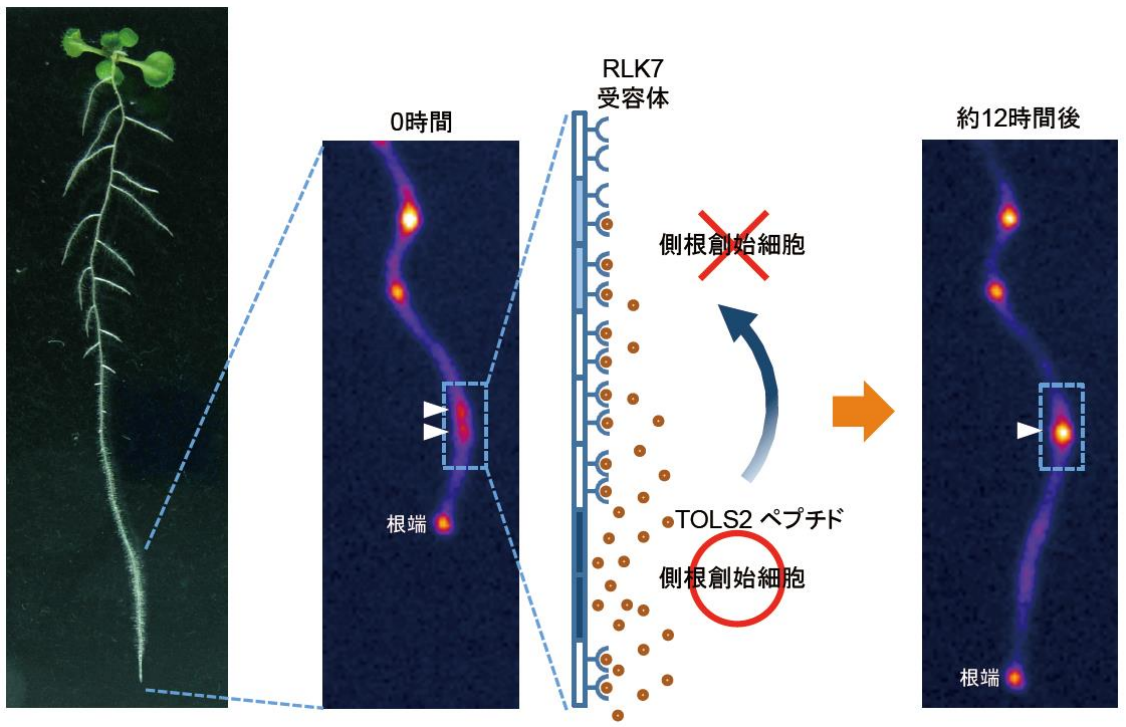
図3 TOLS2ペプチドとRLK7受容体による側根創始細胞の側方抑制
(左写真)シロイヌナズナ野生型(9日目芽生え).(中央と右の写真)根における側根創始細胞のマーカーであるDR5マーカー遺伝子(DR5:Luciferase)の発現を示す。白矢じりは、DR5活性の高い部位。0時間(中央写真)では、DR5活性の高い部位が2つ隣接しているが、約12時間後(右写真)には、片方だけがDR5活性を強め、もう片方は活性を弱める。TOLS2ペプチドとRLK7受容体は、このような側根創始細胞の側方抑制に働くと考えられる。
今後の展開
今後、TOLS2ペプチドによる側根創始細胞の側方抑制の仕組みがシロイヌナズナで明らかにされれば、他の植物種(農作物や樹木)における根系構築の仕組みの理解につながることが期待されます。また、他の植物種にもTOLS2ペプチドと同じ働きをもつペプチドがあれば、それらを利用することで、さまざまな農作物や樹木において、土壌における根系パターンを人為的に制御することが可能になると期待されます。さらに、根系パターンの改良によって植物の成長を調節できるようになれば、農業・林業分野における増産に貢献することも期待されます。
謝辞
本研究は神戸大学を中心に、名古屋大学、奈良先端科学技術大学院大学、大阪大学、東京大学の協力により行われました。また、本研究は以下の研究助成を受けて行われました。
科学研究費新学術領域研究(No. 25110330, 15H05957, 22120006, 16H01234, 25113001)
科学研究費特定領域研究(No. 19060006)
科学研究費基盤研究(S)(No. 25221105, 18H05274)
科学研究費基盤研究(B)(No. 18H02463)
科学研究費若手研究(A)(No. 17H05008)
科学研究費若手研究(B)(No. 25840111)
日本学術振興会特別研究員奨励費 (No. 12J02079, 16J00131)
旭硝子財団研究助成
論文情報
・タイトル
“Lateral Inhibition by a Peptide Hormone-Receptor Cascade during ArabidopsisLateral Root Founder Cell Formation”
https://doi.org/10.1016/j.devcel.2018.11.031
・著者
Koichi Toyokura 1, 2 , Tatsuaki Goh 1, 3 , Hidefumi Shinohara 4 , Akinori Shinoda 1 , Yuki Kondo 5 , Yoshie Okamoto 1 , Takeo Uehara 3, 6 , Koichi Fujimoto 2 , Yoko Okushima 3 , Yoshifumi Ikeyama 3 , Keiji Nakajima 3 , Tetsuro Mimura 1 , Masao Tasaka 3 , Yoshikatsu Matsubayashi 4 , and Hidehiro Fukaki 1
1 神戸大学大学院理学研究科生物学専攻
2 大阪大学大学院理学研究科生物科学専攻
3 奈良先端科学技術大学院大学先端科学研究科
4 名古屋大学大学院理学研究科生物科学専攻
5 東京大学大学院理学系研究科生物科学専攻
6 神戸大学大学院自然科学研究科
・掲載誌
Developmental Cell
参考URL
大阪大学 大学院理学研究科理論生物学教室
http://www.bio.sci.osaka-u.ac.jp/~fujimoto/
用語説明
- 側根創始細胞
将来、側根になる予定の細胞群。シロイヌナズナにおいては、根の原生木部に接する内鞘細胞のうち、根の片側において2列または3列の細胞列で縦に並んだ二つの細胞で、非対称分裂によって側根原基を生じる。
- シロイヌナズナ
アブラナ科シロイヌナズナ属の一年草(学名Arabidopsis thaliana)。2000年に全ゲノム配列が解読され、世代時間が短い、ゲノムサイズが小さい、植物体が小さい、遺伝解析や遺伝子導入が容易などの理由から、モデル植物として幅広く植物研究の材料として利用されている。
- オーキシン
植物ホルモンの一つ。主な天然オーキシンは、インドール-3-酢酸。側根形成だけでなく、植物の胚発生、根・葉・花などの器官形成、果実の発達、光や重力に対する屈性反応など、植物のさまざまな成長・発生、環境応答を調節する。
- ペプチドホルモン
ホルモン様の作用をするペプチド。植物においても、シグナル伝達物質として成長・発生、環境応答に働く。
- DR5
オーキシン応答して隣接する遺伝子の発現を誘導する人工的なDNA配列。DR5配列の制御下で適当なレポータータンパク質を発現させることで、植物体におけるオーキシン応答を可視化することができる。
- CRISPR/Cas9法
部位特異的なヌクレアーゼを用いて標的遺伝子を自在に改変するゲノム編集技術の一つ。