この研究についてひとこと
生物の発生においては、転写因子による細胞タイプの決定と細胞間情報伝達分子、およびこれらの相互調節が鍵を握ります。植物の篩部の形成においてその一端が明らかになりました。今後は、さらに、維管束全体の組織パターンがどのように出来上がるのかを解明したいと考えています。

柿本辰男
- 理学研究科
- 教授
植物の栄養分を運ぶ「篩部」が 正しく作られるためのメカニズム
植物の発生に潜むロマンと大きな可能性
研究成果のポイント
- 篩部(しぶ)は植物の栄養分を運ぶ通路である篩管と、篩管の生命活動を助ける伴細胞からなる。篩部形成のマスター調節因子として篩部の細胞形成の最初のスイッチを入れる転写因子はわかっていなかった。また、篩部になるべき細胞以外は篩部にならないようにしている仕組みもわかっていなかった。
- 篩部領域(篩部前駆細胞も含む)で発現している複数のDofタイプに分類される転写因子(phloem-Dof)は、篩部を構成する細胞になることを指示するとともに、分泌性であるCLEペプチドの合成を指示することを発見。このCLEが周辺細胞に作用して篩部にならないようにしている。
この2つの仕組みが篩部が作られる領域を自発的に決めているということを発見。 - Phloem-Dof転写因子は、篩部細胞形成のマスター調節因子として働くと共に、これらとCLE分泌ペプチドの相互調節によって篩部になる領域を限定していることが解明された。
概要
大阪大学大学院理学研究科のPingping Qian助教、柿本辰男教授らの研究グループは、篩部が形成される細胞で発現する複数のDofタイプ転写因子(phloem-Dof)は篩部の細胞分化を誘導するのみならず、篩部構成細胞の形成を阻害する分泌性ペプチド分子CLE25, 26, 45の合成も誘導することを見出しました。CLE25,26,45はBAM受容体-CIK共受容体複合体によって受容されてphloem-Dofを減少させることにより、本来の篩部形成位置の周りの細胞が篩部にならないようにしていることも見出しました。篩部組織は栄養分を輸送する重要な組織です。篩部形成を制御する因子の報告が相次いでいますが、これらの因子の働きでどのように正しく篩部が作られるのかはわかっていませんでした。
今回の発見で、phloem-Dofは篩部細胞形成制御の最上位で働くこと、転写調節と細胞間のコミュニケーション分子の協調作用が正しい配置で篩部を作り出すことを示したもので、植物の発生の仕組みの一端が明らかになったものと言えます。
本研究成果は、英国科学誌「Nature Plants」に、7月12日(火)午前0時(日本時間)に公開されました。
研究の背景
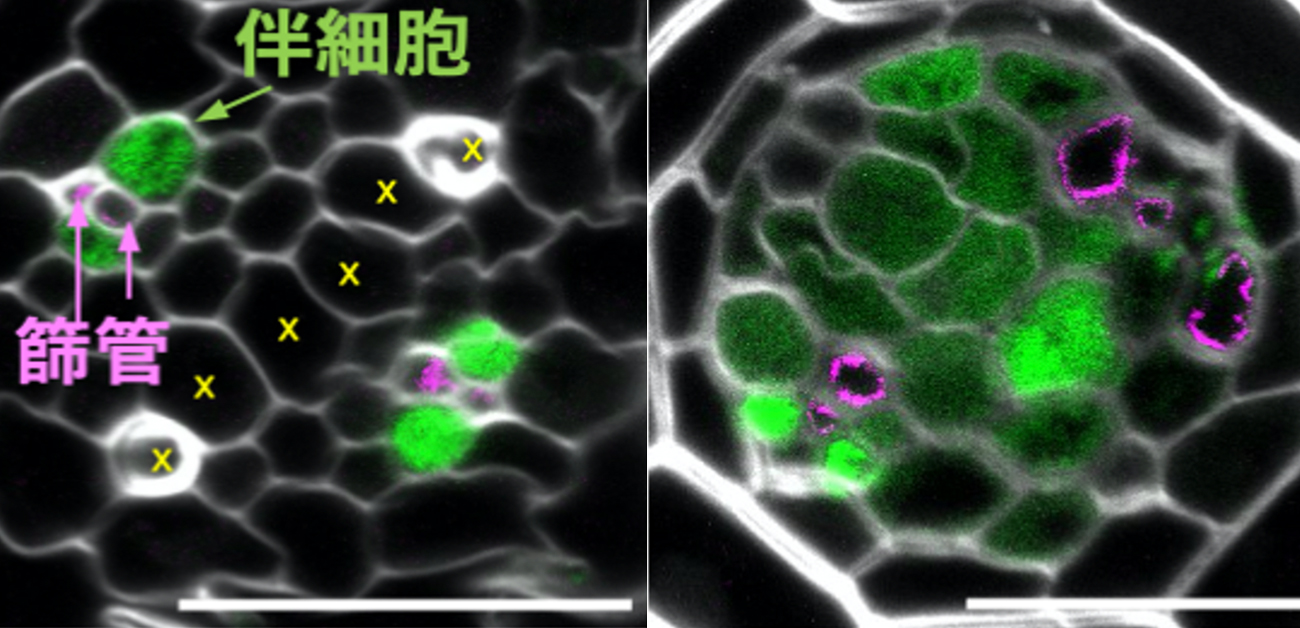
植物の維管束組織は、木部、篩部、前形成層からなります。篩部は、植物の栄養分を運ぶ通路である篩管と、栄養分の積み込み、積みおろしに関わる伴細胞からなっています(図1. 左)。篩管の細胞(篩要素)は生きていますが核を持たず、伴細胞に依存して生きています。木部は道管から構成され、水を輸送します。根では根端近くの細胞は未分化であり、根端から離れるに従って各細胞列の分化が進みます。前形成層は根端から離れても多能性を保っています。
近年、篩部構成細胞の発生の制御系の理解が進んできました。本研究でも重要な位置を占める篩部で発現するDof転写因子が篩部形成の初期過程で重要な役割をしていることもわかってきていましたが、私たちはその役割の理解を深めました。また、CLEペプチドが篩部形成を抑制する作用があることがわかっていましたが、内在のCLEペプチドの機能は、わかっていませんでした。
図1. A.シロイヌナズナの根の断面図。色付けされた部分が維管束で、篩管と伴細胞からなる篩部、道管からなる木部、その間の前形成層からなる。BとC, 篩部で発現している6つのDof遺伝子を破壊すると篩部がほとんどできなくなる。写真部分は全くない。DとE, phloem-Dofの一つDof5.1遺伝子を植物全体で人為的に発現させると本来は篩部ができない位置にも篩部構成細胞が作られる。Dof5.1と配列が似ていて、本来は篩部で発現している他の5つのDofも同様の力を持っていた。逆に、篩部で発現している6つのphloem-Dofの遺伝子を破壊すると篩要素と伴細胞は消失、あるいは極端に減少した。Cにおいて篩部は消失している。
研究の内容
細胞の分化に至る過程では、細胞の性質を決める細胞の転写ネットワークは、少数の転写因子によって決められる例が多いことが知られています。私たちがこの研究を始めた十数年前には、篩部形成を支配する影響力の強い転写因子は知られていませんでした。そこで、篩部の細胞の転写ネットワークの上位に位置し、篩部の細胞を誘導できる転写因子を見つけようと考えました。まず、公開されている細胞種別のマイクロアレーのデータを解析し、篩部で特異的に発現している転写因子を候補として選びました。これらの候補遺伝子を植物全体で発現させたときに本来は篩部ではないところにも篩部の細胞を誘導できる遺伝子を探すという戦略をとりました。篩部の細胞を簡単に見分けられるよう、伴細胞でGFPが発現するSUC2-GFP系統を用い、候補遺伝子を全身的に発現させてGFP 発現への影響を観察しました。このスクリーニングによって、SUC2-GFP発現細胞を誘導できる一群のDofタイプ転写因子(phloem-Dof)を見出しました。また、これらは、篩要素分化の特異的マーカーであるENOD9遺伝子を発現する細胞も生み出し、さらにENOD9発現細胞では篩要素に特徴的な核の消失も起きました。これらのことから、篩部で発現しているphloem-Dofは篩要素と伴細胞を誘導する能力があることがわかります。興味深いことに、SUC2とENOD9が同時に発現する細胞はありませんでした。篩要素になるか伴細胞になるかの運命がどのように分けられるかは今後の課題です。
次に、phloem-Dof遺伝子であるDof1.1, Dof2.2, Dof5.1を植物全体で発現させたときに遺伝子全体の発現パターンがどのように変化するのかをマイクロアレイを用いて解析しました。これらはすべて、篩部形成に関わると知られている遺伝子の発現量を増加させただけでなく、篩部形成を抑制することが知られていたペプチド性シグナル分子CLE25, 26, 45の発現量も増加させました。これらのCLEは通常は篩部で発現しています。そこで、CLE25, 26, 45遺伝子を同時に破壊したところ、篩部形成領域が拡大しました。さらに、これらのCLEの受容体であるBAM1, BAM2, BAM3をコードする遺伝子を破壊した3重変異体や、BAMと共に働く共受容体CIK2とCIK3をコードする遺伝子を破壊しても同じように篩部領域が拡大しました。これらのことから、CLEペプチドが篩部形成の側方阻害因子として働いていることがわかります。
では、CLEによって活性化されたBAM-CIK受容体複合体はどのようにして周辺細胞での篩部形成を抑制しているのでしょうか?私たちは、CLE25がphloem-Dofタンパク質の量を転写後調節によって減少させること(図3)で篩部形成を阻害していることを見出しました。では、篩部になるべき細胞で作られたCLEはその細胞での篩部への分化を抑制しないのでしょうか?phloem-Dofタンパク質はphloem-Dof遺伝子の発現を促進することでポジティブフィードバックを形成しますので、最初に発現を始めた細胞ではDofの発現が安定化します。また、私たちは、phloem-DofはOPSと呼ばれるタンパク質をコードする遺伝子を活性化することも見出しました。OPSはBAMの機能を阻害することが報告されていますので、phloem-DofによるOPSの活性化も、篩部になる予定の細胞でのCLEによる阻害が起きにくくしていると考えられます。この2つの仕組みのために篩部になるべき細胞は篩部になり、周辺細胞が篩部になるのを抑制しているのです。(図4)
図2. CLE25, 26, 45遺伝子を破壊すると、篩部構成細胞が形成される場所が拡張する。この図では、篩管だけにあるタンパク質をマゼンタ色に染めている。CLE25ペプチドを与えると、篩部が形成されなくなる。
図3. CLE25タンパク質はphloem-Dofタンパク質を減少させる。Dof2.4-GFP融合タンパク質を植物全体で発現させた。9本の根のすべての細胞の核に緑色のDof2.4-GFPがあることがわかる(左図)。このような植物にCLE25ペプチドを与えるとDof2.4-GFPが減少する(右)
図4. 篩部前駆体と篩部で発現するDof転写因子は、一群のタンパク質の生成を誘導して篩部の細胞形成を指示すると共に、分泌性シグナル分子CLEの合成を誘導して周辺細胞が篩部の細胞になることを阻害することで、適切に篩部を形成する。
本研究成果が社会に与える影響(本研究成果の意義)
本研究において篩部形成の制御機構が明らかとなり、突然変異や遺伝子操作で篩部になる細胞を増やしたり減らしたりすることができるようになりました。将来は、篩部形成の人為制御によって作物などの栄養分の分布を制御するような技術につながる可能性がないとも言えませんが、それが議論できるような段階ではありません。学術的観点からは、さらに進めて木部、篩部、前形成層の配置が作られる根本原理の解明につながることが期待されます。
特記事項
本研究成果は、2022年7月12日(火)午前0時(日本時間)に英国科学誌「Nature Plants」(オンライン)に掲載されました。
タイトル:“A Dof-CLE circuit controls phloem organization”
著者名:Pingping Qian1 , Wen Song2,4, Miki Zaizen-Iida1, Sawa Kume1, Guodong Wang1,3, Ye Zhang1, Kaori Kinoshita-Tsujimura1, Jijie Chai2,4, and Tatsuo Kakimoto1
所属
1. 大阪大学 大学院理学研究科
2. Innovation Center for Structural Biology, Tsinghua-Peking Joint Center for Life Sciences, School of Life Sciences, Tsinghua University, Beijing 100084, China.
3. College of Life Sciences, Shaanxi Normal University, Xi'an 710119, China
4. Max Planck Institute for Plant Breeding Research, 50829 Cologne, Germany. Institute of Biochemistry, University of Cologne, 50674 Cologne, Germany. Cluster of Excellence in Plant Sciences (CEPLAS), 40225 Düsseldorf, Germany
DOI: https://doi.org/10.1038/s41477-022-01176-0
なお、本研究は、文部科学省日本学術振興会科学研究費助成事業(19H03246, 19K22430, 18H04837, 25113006, L17545,19K23750, 21K15123)、National Natural Science Foundation of China、Alexander von Humboldt Foundation、Max-Planck-Gesellschaftなどの支援により行われました。
参考URL
柿本辰男教授 Researchmap
https://researchmap.jp/read0079052
Qian Pingping Researchmap
https://researchmap.jp/qianpingping2013
SDGsの目標







用語説明
- 篩部
維管束植物の栄養分を運ぶ通路である篩管と、栄養分の積み込みと積み下ろしに関わる伴細胞からなる組織。篩管は篩要素と呼ばれる細胞が小さな穴で縦に連絡していて、篩管液が移動する。篩要素は分化の過程で核を失うが、側方に存在する伴細胞からタンパク質等を送り込まれることにより、生命活動を維持している。
- Dof
植物に特有の一群の転写調節因子。シロイヌナズナには36個存在し、様々な生理機能に関与する。
- 転写因子
DNA配列を認識してゲノムDNAに結合し、標的遺伝子の発現を制御するタンパク質。多くの場合、一つの転写因子が複数の遺伝子を制御する。転写因子により、標的遺伝子の発現を促進するもの、抑制するもの、状況により促進も抑制もするものがある。
- CLEペプチド
植物における分泌性情報伝達ペプチドで、ペプチドごとに様々な生理機能が知られている。遺伝子にコードされたペプチドから12-14アミノ酸残基の成熟型ペプチドとして切り出される。最初に見つかったCLV3はプロリンの一つが糖鎖修飾されることがわかっている。
- BAM受容体
細胞外にロイシンリッチ反復配列からなるシグナル分子受容ドメインを持ち、細胞内にリン酸化酵素ドメインを持つ受容体タンパク質。私たちの発表では、CLEペプチドがBAM3とBAM1の細胞外ドメインに結合する時の親和性も報告している。
- CIK共受容体
受容体よりも小さな細胞外ロイシンリッチ反復配列ドメインと、細胞内セリン・スレオニン リン酸化酵素ドメインから成り、BAM受容体と共に複合体を作っている。CLEを受容すると、CIKとBAMはお互いにリン酸化し合う。
- 転写ネットワーク
違った種類の細胞ごとに、発現している遺伝子の種類と発現量が違っている。細胞種特有の遺伝子発現のパターンがその細胞種のアイデンティティーを決める。この遺伝子発現のパターンは、遺伝子間の相互調節などによって安定化されて、細胞種固有の転写ネットワークを形成する。転写ネットワークを構成する遺伝子の中には影響力が強い遺伝子があり、そのような遺伝子の発現量を人為的に変えることで転写ネットワークと細胞アイデンティティーを変えることができる場合がある。
- 転写後調節
タンパク質の量は、それをコードする遺伝子の転写、mRNAからの翻訳(タンパク質の合成)、タンパク質の分解により調節されている。転写よりも後の調節を転写後の制御と呼ぶ。