

植物の葉緑体に蛋白質を運び入れる分子装置を同定
陸上植物の進化の謎 解明に一歩
リリース概要
大阪大学蛋白質研究所の中井正人准教授および菊地真吾研究員、Jocelyn Bédard研究員らの研究グループは、植物葉緑体の内包膜 から、分子量約100万の新奇な蛋白質膜透過装置 を単離精製する事に成功し、その全構成因子を同定しました。さらに、光産業創成大学院大学の井出徹教授、平野美奈子講師との共同研究で、単離したこの複合体を脂質二重層 に組み込むと、運ばれる葉緑体蛋白質と相互作用する膜チャネルとして機能する事を証明しました。葉緑体は、太古の昔に、光合成をするバクテリアが宿主の細胞に取り込まれる事で誕生しましたが、宿主の細胞内で光合成が効率よく行なわれるためには、葉緑体へ蛋白質を運び入れる仕組みも確立される必要がありました。この役割を担っている分子装置を、植物の葉緑体において同定した今回の発見は、葉緑体の進化や陸上植物の進化に関して新たな知見を与える成果であると同時に、生物が長い進化の過程で生み出してきた数少ない蛋白質膜透過装置のひとつが、本研究により明らかにされたことになります。
研究の背景と今回の成果
藻類や植物の葉緑体は光エネルギーを化学エネルギーに変換する光合成の場であり、それによって作り出される有機物は地球上の多くの生命を支えています。葉緑体は、シアノバクテリア のような酸素発生型の光合成を行なう原核生物 が、10億年ほど前に核やミトコンドリアを持つ真核生物 の一種に細胞内共生 することで誕生したと考えられています( 図1 )。その後、内共生体が持っていた遺伝子の大部分は宿主の核ゲノムへ移行しました。これら核ゲノムにコードされた二千種類を超える葉緑体蛋白質の合成は葉緑体の外(細胞質ゾル)で行なわれるため、葉緑体蛋白質だけを特異的に運び入れる蛋白質膜透過装置が葉緑体を包む膜に同時に進化してくる必要がありました。これまで、葉緑体を包む外側の膜(外包膜)に存在する蛋白質膜透過装置は既に明らかにされていましたが、内側の膜(内包膜)に存在する膜透過装置については良く分かっていませんでした。
今回、中井准教授らは、植物葉緑体内包膜の蛋白質膜透過装置の中核蛋白質を同定し、それに単離精製用のタグ配列を付加したシロイヌナズナの形質転換植物を利用する事で、この膜透過装置を分子量100万の超分子複合体のまま精製する事に世界で初めて成功しました( 図2 )。また、精製した複合体が、運ばれる葉緑体蛋白質と相互作用する膜チャネルとして実際に機能する事を、人工脂質二重層を用いた再構成実験により示しました。
今後の展開
興味深い事に、今回同定された複合体を構成していた4つの蛋白質のうちのひとつは、葉緑体ゲノムにその遺伝子が存在していました( 図2 )。ところが、分子系統解析の結果、この遺伝子は現存のシアノバクテリアや、内共生成立後初期に分岐したと考えられる灰色藻や紅藻には相同遺伝子が見当たらず、したがって、緑藻が出現するまでの進化の過程で葉緑体ゲノムに新たに加わったと考えられる非常に稀な遺伝子である事が分かりました。また、中心蛋白質以外の残りの2つの蛋白質も、緑藻や陸上植物が出現する頃に核ゲノムに遺伝子が加えられた事が分かりました。この事は、シアノバクテリアの内共生による葉緑体の誕生という生物進化上の大イベントの後にも、ダイナミックな遺伝子の水平伝播 などが起き、葉緑体の進化や陸上植物の進化をもたらす一因になった可能性を示唆しています。各蛋白質が加わる事でどのような機能が付加されていったのか、これから明らかにされていくと期待されます。また、このようなダイナミックな変化は継続的に続いており、特に、単子葉植物の中でも比較的最近分岐したイネ科植物では、今回同定された葉緑体内包膜の蛋白質膜透過装置が、中核蛋白質以外は大きく様変わりしている事もゲノム情報から明らかになっています( 図2 )。今後、イネ科で実際にどのような蛋白質膜透過装置が機能しているのかについても、その解明に大きな興味が持たれます。
細胞や細胞内のコンパートメントを取り囲む生体膜は脂質二重層と呼ばれる疎水性のコアを持った分子から成り、そのため、一般に、低分子だけでなく蛋白質などの高分子に対しても、そのままでは通さないバリアとして働きます。しかし細胞が複雑な機能を発揮するためには、これらの分子を、生体膜を隔てて特異的に運ばなければならない場合が数多くあります。特に、蛋白質のような巨大な分子を膜透過させるには複雑で巧妙な仕組みが必要とされます。生命は長い進化の過程で、このような生体膜に組み込まれた蛋白質膜透過装置を、ごく限られた種類、生み出してきた事が分かっています。これまでに、バクテリア型蛋白質膜透過装置であるSEC複合体、TAT複合体、ミトコンドリアのTOMやTIM複合体、ペルオキシソームのPEX複合体などが知られています。しかしこれらの複合体は、互いに、構成蛋白質の組成も違えば、膜透過の分子メカニズムも異なっています。今回発見された葉緑体内包膜の蛋白質膜透過装置も、まったく独自の蛋白質組成を有するもので、生命が生み出してきた巧妙な分子装置のひとつと言えます。今後、今回同定された葉緑体内包膜の膜透過装置による蛋白質膜透過の詳細なメカニズムや立体構造の解明を通して、他の蛋白質膜透過装置との比較が可能となれば、生体膜を隔てて蛋白質を膜透過させるという細胞機能の基本原理のひとつの理解が大きく前進すると予想されます。
本研究成果が社会に与える影響(本研究成果の意義)
植物や藻類の葉緑体が光合成やそれに伴う窒素同化や硫黄同化により生み出す有機物は、地球上の多くの生命活動の源となっています。これら葉緑体の複雑な機能は、2000種類を超える葉緑体蛋白質が葉緑体に正しく運ばれてはじめて発揮されます。今回の発見は、地球上の生命の営みに必須な葉緑体の機能を支える分子メカニズムの解明という意義に加えて( 図3 )、生体膜を隔てた蛋白質分子の膜透過という細胞生物学上の重要な基本原理の理解に結びつくと同時に、葉緑体や植物の進化を扱う学問領域の今後にも大きなインパクトを与えると期待されます。特に、本研究により、イネやトウモロコシ、コムギといった栽培穀物として重要な位置を占めるイネ科植物では、葉緑体への膜透過装置に非常に大きな変化が生じている事が分かり、イネ科植物誕生の謎の解明にも貢献していくものと思われます。
また、今回得られた知見は、将来的には、有用な人工蛋白質や酵素を様々な植物や藻類の葉緑体へ効率よく蓄積させ利用するといった葉緑体工学の分野にも応用が期待されます。
特記事項
本研究成果は、2013年2月1日発行の米国科学誌「Science」に掲載され、それに先立ち、同誌の電子版において( http://www.sciencemag.org/ )、米国東部時間2013年1月31日(木)午後2時(日本時間2月1日(金)午前4時)に公開されます。
論文名および著者名
Uncovering the Protein Translocon at the Chloroplast Inner Envelope Membrane. Science, in press. Shingo Kikuchi, Jocelyn Bédard, Minako Hirano, Yoshino Hirabayashi, Maya Oishi, Midori Imai, Mai Takase, Toru Ide, & Masato Nakai (2013)
参考図

図1
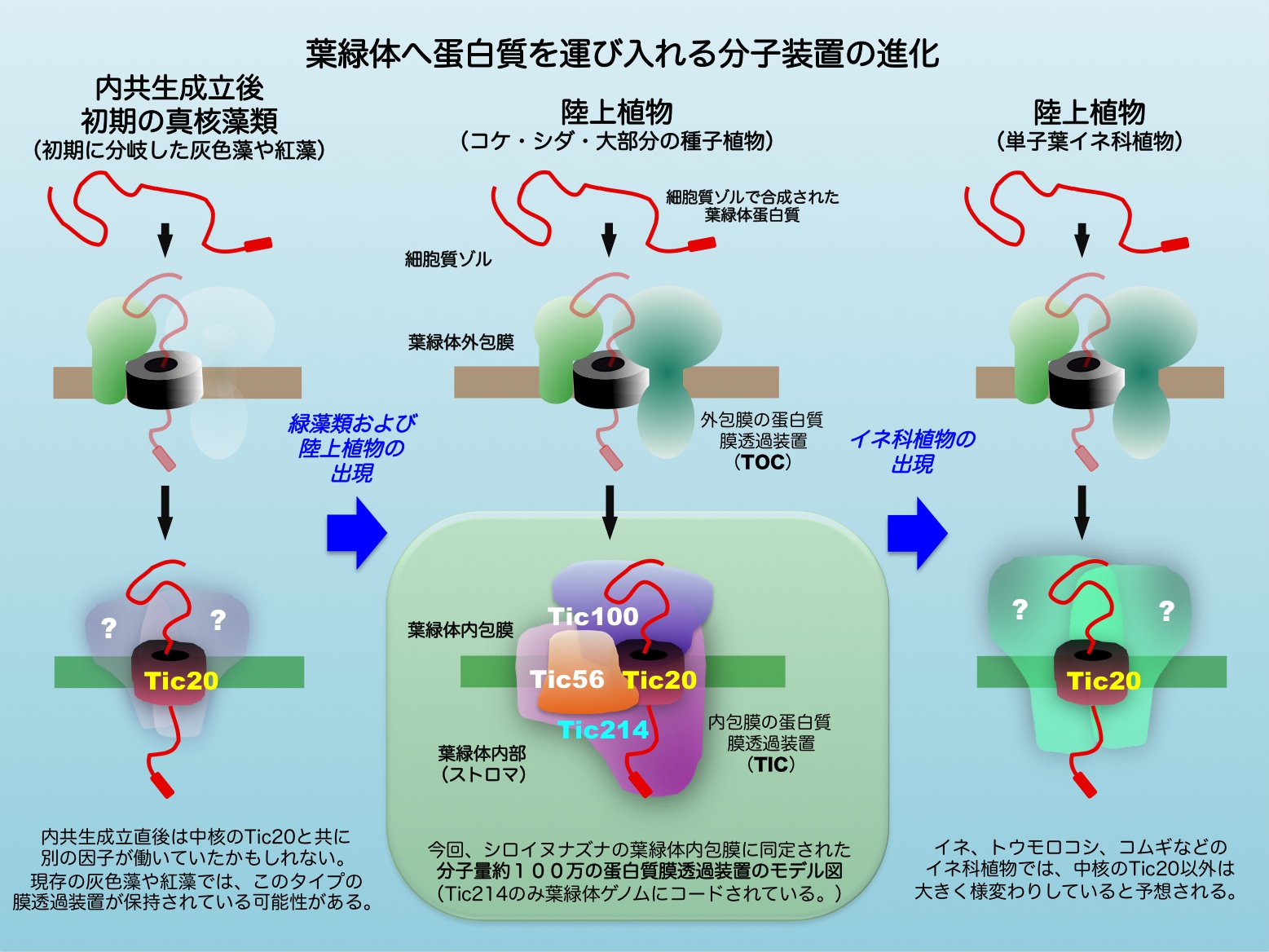
図2

図3 ショ糖を含む寒天培地で2週間育てたシロイヌナズナ。
野生型植物(写真右)が葉緑体が発達した緑色の葉を展開しているのに対し、本研究で同定された膜透過装置に欠陥がある植物体(写真左)では、葉緑体が発達せず、半透明のアルビノとなる。ショ糖を含む培地では、この段階までは育つが、これ以上大きくはならず、花も種もつけない。
参考URL
用語説明
- 葉緑体の内包膜
植物の葉緑体は外包膜および内包膜と呼ばれる二重の包膜に囲まれている。内包膜は、内共生したシアノバクテリアの細胞質膜に由来すると考えられている。光エネルギーをトラップする光化学系が集積するチラコイド膜は、葉緑体の内部に、異なる膜系として存在している。
- 蛋白質膜透過装置
生体膜を隔てて蛋白質を膜透過させるための複合体。通常、一群の蛋白質のみを特異的に膜透過させるために、運ばれる蛋白質が共通に持つシグナル配列を認識するリセプター部分(領域)を有する。一方、膜透過させるための膜チャネル部分は、運ばれる蛋白質の多種多様なアミノ酸配列や二次構造・三次構造を許容する構造的柔軟性を持っていると考えられる。同時に、蛋白質分子のような巨大な分子の通過に伴い、膜本来のシールドとしての役割が失われないような巧妙な仕組みがあると考えられる。また、蛋白質をこのような膜チャネルを通して一方向的に運ぶには、外側から蛋白質をチャネルに送り込む、あるいはチャネル内側から蛋白質を引き込むモーターのような役割をする分子装置が、蛋白質膜透過装置と協調的に働く必要がある。蛋白質膜透過は細胞の機能発現と秩序維持の根幹に関わる必須の仕組みである。 (参考:特定領域研究「タンパク質の社会」 http://www.protein.bio.titech.ac.jp/outline.html )
- 脂質二重層
生体膜を構成する基本構造。脂肪酸からなる疎水性コアと表面に露出したリン酸基などの親水性部分からなる。イオン等の低分子や蛋白質などの高分子は通る事ができない。細胞の内と外とを隔てる他、ミトコンドリアや葉緑体といった細胞内の小器官も生体膜によって区画化されている。
- シアノバクテリア
30億年ほど前に出現した酸素発生型の光合成を行なう原核生物。シアノバクテリアの繁栄により地球に分子上酸素が蓄積した。細胞内に光合成装置(光化学系)複合体を集積したチラコイド膜を持つ。
- 原核生物
細胞核を持たない生物。原始生命体として最初に地球上に出現した細胞の形態と考えられる。真正細菌と古細菌に分類され、シアノバクテリアは真正細菌に属する。これに対し、藻類や動植物、カビやキノコといった菌類など、細胞内に遺伝情報であるDNAを含む核と呼ばれる構造を有する生物を真核生物という。
- 細胞内共生
ある生物の細胞内に別の生物・細胞が共生する事。現在でも様々な形態の細胞内共生が観察される。通常、代謝産物の相互利用という点で「持ちつ持たれつ」の関係になっている。共生関係を無くしても宿主や共生体が生存する場合もある。細胞内共生が長い年月をかけて進化を遂げる事により、ミトコンドリアや葉緑体といった真核細胞の細胞内小器官が生じたと考えられる。真核藻類の葉緑体がシアノバクテリアの一次共生から生じたとすれば、真核藻類が共生したと考えられる二次共生生物、さらにそのような二次共生生物が共生したと考えられる三次共生生物なども存在している。このような複雑な共生関係が劇的な生物進化の原動力のひとつとなってきたと考えられる。 (参考:新学術領域研究「マトリョーシカ型進化原理」 http://www.matryoshka-evolution.jp )
- 遺伝子の水平伝播
生物は通常、遺伝により遺伝子を受け継ぐ。この際に低頻度で生じる突然変異や遺伝子重複や組換えが進化の痕跡として、時間軸で言えば垂直方向で蓄積していく。ところが、近年、多くの生物で、このような単純な遺伝では説明できない遺伝情報の変化が進化の過程で度々起きている事が分かっている。その多くは、ウイルスや寄生などを介して他の生物の遺伝子が取り込まれた事によると説明できる。このような時間軸において同時期の、すなわち水平方向の遺伝子の伝播の事を「遺伝子の水平伝播(HGT: Horizontal gene transferもしくは LGT: Lateral gene transfer)」という。内共生体の遺伝子が核へと転移していく現象は特に EGT: Endosymbiotic gene transfer と呼ばれる。葉緑体やミトコンドリアが持っていたゲノムからの核への遺伝子の転移はEGTにあたり、これは現在も継続的に続いている。