
走化性細胞が応答範囲を拡張するメカニズム
生物が環境適応する仕組みの一端を解明
概要
理化学研究所(理研)生命システム研究センター細胞シグナル動態研究グループの上田昌宏グループディレクター(大阪大学大学院生命機能研究科 教授(研究当時:理学研究科))らの研究チーム は、「走化性」における応答範囲を調節する因子「Gip1」を発見しました。この因子は、三量体Gタンパク質 の細胞内局在制御というこれまで知られていなかったメカニズムで、走化性の応答範囲を拡張していることが分かりました。
細胞は化学物質の濃度勾配に沿って移動することができます。このような現象は「走化性」と呼ばれ、胚発生、免疫、神経回路形成、傷口の治癒などに重要な役割を果たしています。走化性細胞は10万倍にも及ぶ広い化学物質の濃度範囲にわたって、数%程度の微小な濃度差を認識できます。このような性質は生物の持つ柔軟な環境適応の仕組みとして、古くから注目を集めてきました。
今回、研究グループは、走化性応答の濃度範囲を広げるタンパク質Gip1を同定しました。この因子は細胞膜上での三量体Gタンパク質の量を調節し、受容体からのシグナルを適正に伝達していました。Gip1は三量体Gタンパク質との結合を介してその一部を細胞質画分として保持し、外界の化学物質の増加に応じて細胞質内の三量体Gタンパク質を細胞膜へ送り出していました。このような三量体Gタンパク質の局在調節という新たなメカニズムにより、走化性細胞は適切に濃度勾配を認識し、応答範囲を拡張していました。
走化性細胞を含め多くの感覚細胞は、広範な刺激に対して応答できます。このような応答範囲の拡張メカニズムとして、受容体の化学修飾による制御がこれまで広く知られていました。今回の研究では三量体Gタンパク質の局在制御という新しいメカニズムも、その調節に関わっていることを突き止めました。このような制御は真核生物で広く保存されている可能性があり、三量体Gタンパク質がGip1との結合により細胞質にとどめられる仕組みなど、その分子基盤に関する知見が今後、他のシステムでの理解に貢献すると期待できます。
本研究は、米国の科学雑誌『Proceedings of National Academy of Science of the United States of America (PNAS)』に掲載されるのに先立ち、オンライン版(4月5日付け)に掲載されました。
※ 研究チーム
・理化学研究所 生命システム研究センター 細胞シグナル動態研究グループ
グループディレクター 上田昌宏(うえだまさひろ)(大阪大学大学院 生命機能研究科 教授(研究当時:理学研究科))
上級研究員 上村陽一郎(かみむらよういちろう)
・大阪大学大学院 理学研究科
助教 宮永之寛(みやながゆきひろ)(大阪大学大学院 生命機能研究科 助教)
研究の背景
細胞は、化学物質の濃度勾配に沿って移動することができます。このような現象は「走化性」と呼ばれ、胚発生、免疫、神経回路形成、傷口の治癒などに重要な機能を果たします。走化性の研究には土壌微生物である細胞性粘菌 (キイロタマホコリカビ、 Dictyostelium discoideum )が用いられてきました。粘菌細胞は通常バクテリアを餌として生育していますが、栄養飢餓になると自らが産生、分泌する環状アデノシン一リン酸(cAMP) という化学物質に対して走化性を示すようになります。走化性物質cAMPは細胞膜上のGタンパク質共役型受容体(GPCR) に結合し、三量体Gタンパク質によって下流にシグナルを伝達します。そして運動装置のアクチン骨格系の制御を通して、最終的に細胞は方向性のある運動に至ります。このような分子基盤は、多くの走化性細胞においても保存されています。
分子基盤の共通性に加え、走化性細胞は一般的に、10万倍にも及ぶ広い濃度範囲にわたって数%程度の微小な濃度差を認識できます。このような広範囲にわたる応答は他の感覚細胞でもみられるもので、例えば、バクテリアの走化性や動物の視覚系では、受容体の化学修飾を介した制御を軸に、その理解が進んできました。粘菌細胞でも受容体のリン酸化が起こらないと、走化性における濃度範囲は狭くなるものの、その影響は限定的でした。このような事実は、まだ知られていない制御機構の存在を示唆するものでした。
研究手法と成果
研究チームは粘菌細胞を用いて、三量体Gタンパク質と相互作用し、走化性を制御する因子を探しました。その結果、既知の結合タンパク質とは構造的に相同性のない因子を同定し、Gip1(trimeric G protein i nteracting p rotein 1 )と名付けました。Gip1はPHドメイン と、ヒトTNFAIP8 と弱い類似性を持つ領域から構成されており、後者の領域で三量体Gタンパク質と結合していることが生化学的解析から明らかになりました。この複合体は、三量体Gタンパク質のβγサブユニットを介して形成されていました。
次に、Gip1と走化性との関係を調べるため、粘菌細胞の gip1 遺伝子破壊株 を作製しました。 gip1 破壊株は、高濃度の走化性物質に対する走化性能が極端に低下するという表現型を示しました (図1A、B) 。一方、細胞内のGip1量を増やすと、低濃度での走化性があまり起こりませんでした (図1B) 。つまり、Gip1は走化性における濃度範囲の調節に関わっていることが分かりました。

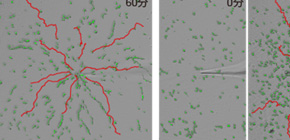
図1 粘菌細胞における野生型、 gip1 破壊細胞の走化性能の比較
A: 粘菌細胞に、マイクロピペットから走化性物質であるcAMPを拡散し濃度勾配を与えた。60分後の野生株および150分後の gip1 破壊株の細胞の軌跡を赤線で示している。細胞は緑色で表示。野生株はピペットの先端まで濃度勾配をさかのぼることができるが、 gip1 破壊株は途中で止まっている。つまり、cAMP濃度が高いマイクロピペットの先端付近で、 gip1 破壊株は走化性能を失っていることを示している。
B: さまざまな濃度のcAMPに対する細胞の走化性能の検定をした。野生株(薄緑線)に比べて、 gip1 破壊株(ピンク線)では高濃度で走化性能が低下しており、Gip1過剰発現株(紫線)では低濃度で走化性能が低下している。
それでは、Gip1はどのように走化性の濃度範囲を調節しているのでしょうか。Gip1は三量体Gタンパク質と結合することから、粘菌細胞の三量体Gタンパク質の活性化あるいは不活性化の反応を調べてみましたが、野生株と比べて大きな違いはなく、正常に機能していました。このことは、Gip1による制御がこれまでに知られていないメカニズムによることを示唆するものでした。
そこで、研究チームは三量体Gタンパク質の細胞内局在に着目しました。通常、三量体Gタンパク質は細胞膜上に局在して機能しますが、その一部は細胞質にもあることが知られていました。しかし、この細胞質中の三量体Gタンパク質が重要であるかは不明でした。研究チームは gip1 破壊株で、細胞質中の三量体Gタンパク質の量が著しく低下していること、逆に細胞内でのGip1量を増やすと、細胞膜上の三量体Gタンパク質が減り、そのほとんどが細胞質に移行することを見出しました (図2) 。Gip1の細胞質局在を考慮すると、Gip1は三量体Gタンパク質と細胞質で結合し、Gタンパク質の一部を細胞質にとどめる機能を持つことを示していました。つまり、三量体Gタンパク質はGip1を介して細胞膜と細胞質を行き来(シャトリング)していることが明らかになりました。

図2 粘菌細胞における三量体Gタンパク質の細胞内局在
A: 三量体Gタンパク質の細胞内局在を調べるため、Gタンパク質のαサブユニットを蛍光標識し観察した。白くみえる部分がαサブユニットのシグナルである。野生株では細胞膜と細胞質の両方にシグナルがみられるが、 gip1 破壊株では細胞質のシグナルが低下している。また、Gip1を過剰発現した細胞では細胞膜でのシグナルが大きく減少している。
B, C: 三量体Gタンパク質の細胞内局在を、生化学的に細胞成分を分けることで検証した。Bは、細胞(W)を細胞質(S)と細胞膜(P)に分け、ここに含まれるGタンパク質のαサブユニット量を抗体により可視化したもの。Cは、細胞質画分のGタンパク質の相対量を示すグラフ。野生株(wt)では約40%のαサブユニットが細胞質に局在するが、 gip1 破壊株( gip1 △)では約10%に減少し、Gip1過剰発現株(Gip1 OE )では約80%にまで上昇している。
さらに、一連の実験から三量体Gタンパク質のシャトリングが走化性において重要な役割を果たすことが明らかになりました。三量体Gタンパク質は走化性物質の刺激により、細胞質から細胞膜へと局在を変えてその量が調整され (図3A) 、濃度勾配に応じて細胞膜上に空間的な偏りを形成していました (図3B) 。このような制御により、細胞は走化性物質の濃度が高くなっても細胞膜上で受容体からのシグナルを適切に伝達し、濃度勾配の方向を識別していました (図3C) 。また、三量体Gタンパク質の局在制御には、Gip1のPHドメインに作用する未知のシグナルが関与することが示唆されました。
このような理解は、Gip1に関連する走化性の表現型を矛盾なく説明できます。例えば、野生型では走化性物質の濃度が高くなるにつれ、細胞質の三量体Gタンパク質が細胞膜へと供給され、十分な勾配シグナルを送ることができます。一方、 gip1 破壊株では三量体Gタンパク質のシャトリング機構がないため、高濃度において細胞質から細胞膜への再配分が起こらず、濃度勾配を認識できません。また、Gip1を大量に発現すると、細胞膜上の三量体Gタンパク質が枯渇し、特に走化性物質の低濃度領域では十分なシグナルを生み出せませんが、濃度が上がるにつれ三量体Gタンパク質の膜移行が起こり、濃度勾配を正常に感知できるようになります。
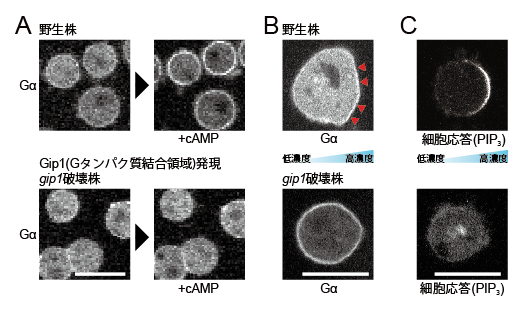
図3 粘菌細胞における走化性物質による三量体Gタンパク質の細胞内局在制御
A: cAMP刺激の前後で蛍光標識したGタンパク質のαサブユニットを観察した。白くみえる部分がGタンパク質のαサブユニットのシグナルである。野生株では、刺激により細胞質から細胞膜へ局在が変化する。 gip1 破壊株は、Gip1の三量体Gタンパク質結合領域だけを発現させても局在変化は起こらない。
B: cAMP勾配中でのGタンパク質αサブユニットを観察した。野生株では高濃度側(赤い▲部分)の細胞膜上に局在するが、 gip1 破壊株では均一に局在したままである。
C: 細胞の応答を調べるために、細胞極性に沿って局在する脂質分子(PIP 3 )に特異的に結合する分子を蛍光標識し観察した。野生株ではPIP 3 の偏りができるが、 gip1 破壊株では走化性物質の勾配情報を適切に処理できず、PIP 3 の偏りが観察されない。
本研究から、研究チームは 図4 のようなモデルを提案しています。
(1)Gip1は三量体Gタンパク質と結合し、細胞質プールを作る。
(2)この細胞質プールは走化性物質依存的に細胞膜へ移行する。
(3)外界の走化性物質の濃度勾配に応じて、細胞膜上の三量体Gタンパク質の局在の勾配を作る。
このように三量体Gタンパク質が、細胞膜上へ適切に供給されることで、走化性細胞は濃度勾配を認識することができるようになります。今回明らかになった三量体Gタンパク質の局在調節による応答範囲の拡張は、広く知られている受容体の化学修飾による制御とは異なる新しい調節メカニズムです。
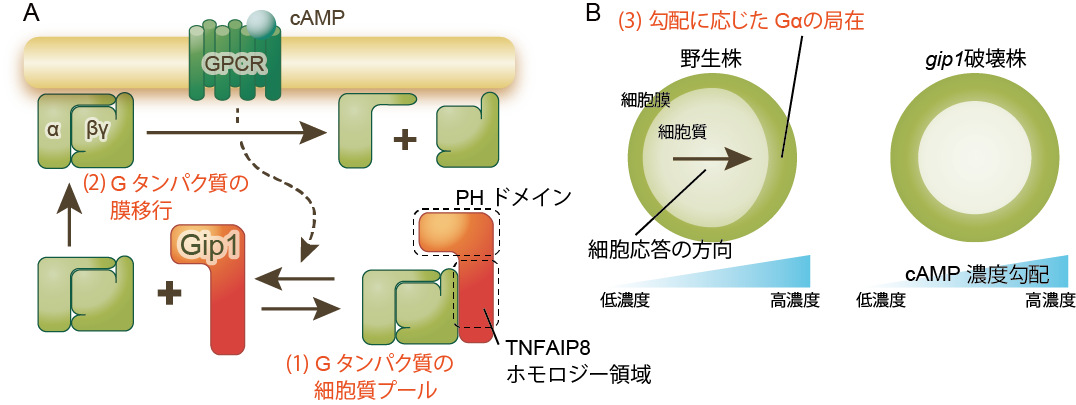
図4 走化性における濃度範囲拡張のメカニズム
A: Gip1タンパク質は三量体Gタンパク質と結合することで、その一部を細胞質にとどめている。走化性物質cAMPがGタンパク質受容体(GPCR)に結合すると、細胞質プールの三量体Gタンパク質がGip1から解離し、細胞膜へと移動する。
B: 野生株は、走化性物質cAMPの濃度勾配下で三量体Gタンパク質(αサブユニット)もそれに応じた偏りを示すが、 gip1 破壊株では細胞膜上の局在は均一なままである。このことは、細胞質の三量体Gタンパク質が細胞膜上で再配置することで、濃度範囲拡張に効いていることを示している。
今後の期待
今回の研究から、粘菌細胞におけるタンパク質Gip1を介した三量体Gタンパク質のダイナミックな局在制御が明らかになりました。このような制御は真核生物で広く保存されている可能性があり、三量体Gタンパク質がGip1との結合により細胞質にとどめられる仕組みなど、その分子基盤に関する知見が他のシステムでの理解に貢献すると期待できます。また、三量体Gタンパク質の細胞膜移行には未知のシグナルが関わっていました。今後、走化性シグナル伝達の理解には、この経路を明らかにすることが不可欠になります。さらに、この応答にはGip1のPHドメインが関与しますが、その機能は分かっていません。一般的に、PHドメインは細胞膜で働くタンパク質に重要であると考えられていますが、Gip1は細胞質で機能します。このことから、今後の解析によって、これまでに考えられていなかったPHドメインの新しい機能の理解に結びつく可能性があります。
三量体Gタンパク質の局在制御は、走化性における勾配認識に関与していました。今回の研究から走化性物質の濃度が低い場合、細胞膜上にある三量体Gタンパク質により勾配シグナルが伝達されますが、高濃度になると三量体Gタンパク質の局在変化による細胞膜上での空間的な偏りが必要になることが分かりました。今後は、三量体Gタンパク質の活性化と局在制御を組み込んだ数理モデルの構築により、広い濃度範囲において濃度勾配情報が受容・処理・伝達される仕組みの理論的な理解が可能になると期待されます。
GPCR-三量体Gタンパク質は、真核生物において最も広く使われているシグナル分子であり、ヒトにおいてはGPCRの多くが疾病治療薬のターゲットになっています。今回の成果は、三量体Gタンパク質の局在制御という新しい視点でGPCRのシグナル伝達を操作・変調しうる可能性を示しています。
論文情報
<タイトル>
Heterotrimeric G protein shuttling via Gip1 extends the dynamic range of eukaryotic chemotaxis
<著者名>
Yoichiro Kamimura,Yukihiro Miyanaga,Masahiro Ueda
<雑誌>
Proceedings of National Academy of Science of the United States of America
<DOI>
10.1073/pnas.1516767113
参考URL
大阪大学大学院生命機能研究科・1分子生物学研究室
理化学研究所・生命システム研究センター・細胞シグナル動態研究グループ
上田グループ研究室HP
http://www.bio.sci.osaka-u.ac.jp/bio_web/lab_page/ueda/
用語説明
- 三量体Gタンパク質
Gタンパク質共役型受容体(GPCR)からの情報を伝えるタンパク質複合体で、アルファ(α)、ベータ(β)、ガンマ(γ)の3つのサブユニットから構成されている。αサブユニットは、グアノシン二リン酸(GDP)あるいはグアノシン三リン酸(GTP)と結合することで不活性型と活性化型の切り換えが起こる。また、βとγは強固な複合体を作り、ひとまとまりとして機能する。つまり、Gタンパク質はGDP結合型αサブユニットがβγサブユニットと三量体を形成し不活性化されているが、GPCRによって活性化されると、αサブユニットのGDPはGTPに置き換わり、βγサブユニットから解離する。そして、αあるいはβγサブユニットが下流へとシグナルを伝える。
- 細胞性粘菌
細胞性粘菌は土壌微生物であり、このうちキイロタマホコリカビ( Dictyostelium discoideum )はモデル生物として広く利用されている。その全ゲノムは2005年に解読され、遺伝子破壊など分子遺伝学的解析や液体培地での大量培養なども容易である。土壌中では単細胞としてバクテリアを餌に増殖しているが、栄養が枯渇すると約10万個の細胞が集合し、多細胞体化する。この多細胞体は一連の形態変化を経て、胞子を含む子実体を形成する。キイロタマホコリカビは細胞運動、分化の研究モデルとしてだけでなく、創薬資源としても注目されている。
- 環状アデノシン一リン酸(cAMP)
アデノシン三リン酸からアデニル酸シクラーゼという酵素によって合成される分子。リボースの3’と5’のリン酸が環状に結合している。多くの細胞で二次伝達物質として多様な生理機能に関与している。細胞性粘菌では、走化性物質としても利用されている。
- Gタンパク質共役型受容体(GPCR)
真核生物で最大のファミリーを形成する膜タンパク質受容体。GPCRはさまざまな刺激を受け、細胞膜上で三量体Gタンパク質を介してシグナルを伝達する。このため、Gタンパク質共役型受容体と呼ばれている。また、共通の構造として、7つの膜貫通領域を持つことから7回膜貫通型受容体ともいわれる。
- PHドメイン
血小板におけるプロテインキナーゼCの基質であるプレクストリンタンパク質中に見出された約100個のアミノ酸からなるドメイン。ドメインとは、タンパク質中にある構造上保存された領域のこと。このドメインには、ホスファチジルイノシトールポリリン酸が結合することが知られている。PHは、 p leckstrin h omologyの略。
- TNFAIP8
ヒトがん細胞で比較的高いレベルで発現するタンパク質として同定された。ヒトにはTNFAIP8と相同性のあるタンパク質がほかに3つあり、がんや免疫などへの関与が示唆されている。TNFAIP8は、 t umor n ecrosis f actor- a lpha- i nduced p rotein 8 の略称。
- 遺伝子破壊株
遺伝子破壊は、ある遺伝子の機能を調べる際に用いられる遺伝学的手法の1つで、これによって作製された細胞を遺伝子破壊株という。細胞性粘菌の場合、遺伝子の翻訳領域に、例えば薬剤耐性遺伝子発現カセットを挿入したものを細胞内に導入し、相同性組換えによってゲノム内の遺伝子を破壊することができる。