

哺乳類初期胚の膜ダイナミクス
胚への栄養供給・分化シグナルを制御するミクロオートファジーの機能解明
リリース概要
大阪大学産業科学研究所の和田洋准教授、大学院医学系研究科の原田彰宏教授、同志社女子大学薬学部の川村暢幸助教、和田戈虹教授、秋田大学大学院医学系研究科の高須賀俊輔助教、佐々木雄彦教授らの研究グループは、マウスの初期発生胚で、ミクロオートファジーとよばれるユニークな膜ダイナミクスが起きていること、ミクロオートファジーにはrab7タンパク質の機能が必要であることを明らかにしました。rab7の機能を喪失したマウス胚は、胚組織の形成が正常に進行しないため受精後6日目以降の形態形成が停止します。一連の研究により、胚が物質・栄養を獲得して、増殖と分化が協調して進行する際、ミクロオートファジーを積極的に利用していることを明らかにし、母体内で受精から形態形成が起きる哺乳動物での母子間相互作用に新たな視点をもたらしました。
本研究成果は英国の科学雑誌「Nature Communications」で9月18日(英国ロンドン時間)公開されます。
背景
我々ヒトに最も近いモデル生物であるマウスにおいて、受精卵は細胞の数を増やすと同時に細胞を分化させ、8日目までに内胚葉、中胚葉、外胚葉とよばれる基本的な組織を構築します。この増殖と分化には大量に物資とエネルギーが必要です。胎盤が形成される以前のこの時期、母体から供給される免疫グロブリン やトランスフェリン などの栄養物質は、胚の細胞の表面に結合し、細胞表面の膜の一部とともに細胞内に陥入して、小さな袋(膜小胞)となって細胞内へ運び込まれます。膜小胞は細胞内のエンドソーム・リソソーム とよばれる細胞内小器官に運ばれます。この一連の過程はエンドサイトーシス とよばれており、初期発生胚の細胞に限らず、様々な細胞で起きることが知られています。
栄養の吸収に重要な臓側内胚葉とよばれる細胞グループでは極めて活発にエンドサイトーシスが起きており、細胞内小器官が発達しています。細胞表層、細胞内の膜小器官同士は互いに物質と情報をやりとりしてダイナミックなネットワークを形成しています。エンドサイトーシスは細胞の増殖や分化、さらには細胞同士の位置関係を支配する情報伝達システムにも深く関わっています。短時間の間に飛躍的に細胞の数が増え、相互作用しながらかたちづくりが進んでいく初期発生では栄養の供給、細胞間の情報伝達はきわめて重要な意義をもっています。しかし、初期発生胚におけるエンドサイトーシスは未解明な点が多く残されています。
研究の内容
蛍光色素を結合させた高分子物質を初期発生胚に取り込ませ、時間を追って細胞内のどこに運搬されるのか観察し、エンドサイトーシスの終着点が頂端液胞 という巨大な空胞であることがわかりました。通常、膜で構成される小胞同士がひとつの空胞になる場合、小胞同士が接着し、互いの膜が融合して一つの連続した膜をつくる、膜融合を伴う小胞融合が主なメカニズムです(図:膜融合を伴う小胞融合)。しかし、初期発生胚の空胞では、大きな空胞が小さな前駆体を飲み込んだ後、空胞内で膜を分解してひとつの小器官になることがわかりました。この膜のダイナミクスは微小自食作用(ミクロオートファジー )としてしられています(図:ミクロオートファジーによる小胞融合)。哺乳動物細胞でのミクロオートファジーはほとんど研究がなされておらず、初期発生胚で極めて活発なミクロオートファジーが起きていることは新しい発見です。
さらに、空胞の膜ダイナミクスに重要なタンパク質、rab7を欠失したマウス胚ではミクロオートファジーが起きず、非常に小さな断片化した空胞が蓄積するものの、巨大な頂端液胞は形成されないことがわかりました。ミクロオートファジーに欠損をもつマウス胚は外胚葉と内胚葉の分化と形成がおきます。しかし、中胚葉組織が低形成となり、受精後6日目以降の発生が停止してしまいます。すなわち、ミクロオートファジーによる細胞内の物質輸送は、初期発生にきわめて重要であることがわかりました。
今後の展開
初期発生胚のエンドサイトーシスでは、エンドソームがまるごとリソソームに取り込まれる、ミクロオートファジーが起きること、また、ミクロオートファジーの欠損が着床直後の初期発生の停止につながることを見出しました。今後、発生の停止が貧栄養に由来するのか、あるいは情報伝達システムの破綻に由来するのか、あるいは複合的な結果であるのかをあきらかにすることが必要と考えます。進化の過程で哺乳動物が獲得した新しいストラテジー:母体内での発生-を支える細胞機能としてエンドサイトーシスに注目することにより、受胎のメカニズムに新しい知見を得ることが期待できます。
特記事項
本成果は、文部科学省 科学研究費補助金 新学術領域研究「哺乳類初期発生の細胞コミュニティー」(領域代表者: 藤森俊彦 基礎生物学研究所 教授)などによって得られました。
参考図
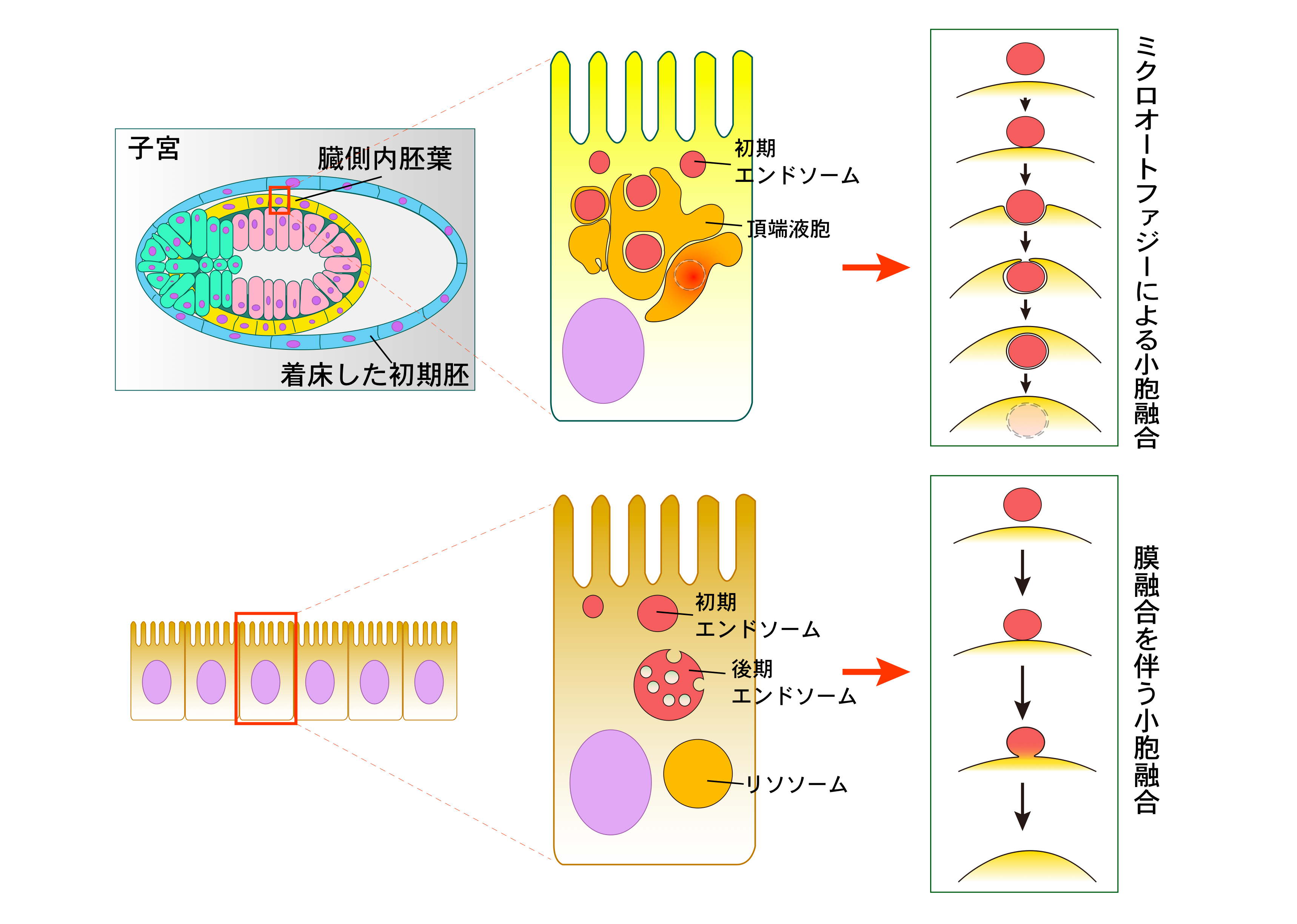
ひとつの細胞であった受精卵は分裂によって数を増やしながら、子宮に着床し、受精後5-6日目に臓側内胚葉とよばれる一層の細胞層(黄色)を分化させる。壁側内胚葉(水色)の層は細胞がまばらであり、子宮からの栄養物質は通り抜けることができる。しかし、臓側内胚葉は細胞が密で、細胞の中を通らなければ、胚本体(ピンク)に届かない。
臓側内胚葉の細胞はエンドサイトーシスによって細胞内の栄養を取り込み、胚本体に物質とエネルギーを供給している。これを反映して、臓側内胚葉は細胞質に大きな液胞(頂端液胞)を発達させて活発なエンドサイトーシスを支えている。エンドソームが液胞に融合する際、頂端液胞がエンドソーム全体を飲み込み、そのあと、エンドソームを液胞内で分解する、ミクロオートファジーが起きている(左図上参照)。
これまで知られているエンドソームとリソソームの相互作用には、エンドソーム膜と頂端液胞膜が直接、単一の膜を形成する膜融合を伴うとされている(左図下参照)。ミクロオートファジーでは頂端液胞の膜面積をむやみに増加させることなく、むしろ、減少させる機構であり、活発なエンドサイトーシスによる頂端液胞の肥大化を抑制している可能性が考えられる。
参考URL
用語説明
- ミクロオートファジー
細胞内のタンパク質や構造を分解するメカニズムのひとつとしてオートファジー(自食作用)がしられている。ミクロオートファジーは、リソソームの膜が陥入して細胞内構造体や細胞基質を直接、リソソーム内に運び込む過程を指す。
- ab7タンパク質
低分子量GTP結合タンパク質のひとつで、細胞内の膜コンパートメント、エンドソームとリソソームの間の物質輸送を制御している。シャルコー・マリー・トゥース病として知られる神経変性疾患の原因遺伝子であることが明らかになっている。
- 免疫グロブリン
血液や体液に含まれるタンパク質のひとつで、抗原に特異的に結合して排除するのに機能している。
- トランスフェリン
血液に含まれるタンパク質のひとつで鉄イオンを結合して体中に運搬する役割をもつ。
- エンドソーム・リソソーム
細胞の中に存在する、膜に囲まれた小さな袋状(小胞)の構造体。細胞膜の一部が細胞内にくびれ込み、小胞を形成する。小胞は細胞内を移動しながら性質が徐々に変化し、初期エンドソーム・後期エンドソームを経て、最終的にはリソソームとなる。細胞外のトランスフェリンや免疫グロブリンは細胞膜の受容体とともにエンドソームに取り込まれる。
- エンドサイトーシス
エンドソームを経て細胞外の物質が細胞内に取り込まれる過程をエンドサイトーシスとよぶ。
- 頂端液胞
植物細胞では、リソソームに相当する構造体が非常に大きく発達しており、液胞(vacuole)とよばれている。一方、動物細胞では通常、リソソームはそれほど大きな構造にはならないが、初期胚の内胚葉細胞や、空腸上皮細胞にはたいへん大きなリソソームがつくられることが知られている。この大きなリソソームは上皮細胞の頂端側(apical)にあることから、頂端液胞(apical vacuole)とよばれる。