この研究についてひとこと
小胞体とミトコンドリアは仲の良い隣人同士のような関係を築いており、ときには連携をとりながら機能を果たすことが報告されてきましたが、今回、小胞体のタンパク質がミトコンドリアを「壊す」マイトファジーに貢献する機構の一端を明らかにすることができました。小胞体にいながらマイトファジーの制御に関わる因子が他にも存在するのか、そうだとすればどういったメカニズムで関わるのか、今後の解析が待たれます。(大西真駿 特任研究員(当時))
岡本浩二
- 生命機能研究科
- 准教授
ミトコンドリア の健康管理機構「マイトファジー」には小胞体との連携が重要!
タンパク質配送システムの新たな役割を解明
研究成果のポイント
- これまでの研究から、ミトコンドリアを丸ごと分別・除去する仕組み「マイトファジー」の進行には、小胞体への膜タンパク質挿入を行うGET経路が関与することが見出されていたが、Get因子がどのようにミトコンドリアの分解に寄与するかは未解明だった
- 今回、ミトコンドリアの分解機構に、GET経路と、ミトコンドリアの膜タンパク質排出装置Msp1が関わること、およびそれらの仕組みの一端を解明
- GET経路は、マイトファジーの抑制因子Ppg1-Far複合体を小胞体膜に繋ぎ止め、ミトコンドリアへの過剰な局在化を防ぐことで、マイトファジーの駆動に寄与していた
- ミトコンドリア外膜タンパク質Msp1は、ミトコンドリアへと配送されたPpg1-Far複合体を除去し、マイトファジーの進行に働く可能性が示された
概要
大阪大学大学院生命機能研究科の大西真駿特任研究員(研究当時、現・マックスプランク老化生物学研究所ポストドクトラルフェロー)、岡本浩二准教授の研究グループは、オートファジーを介したミトコンドリアの分解機構に、小胞体の膜タンパク質挿入装置GET経路と、ミトコンドリアの膜タンパク質排出装置Msp1(図1)が関わることを明らかにしました。
ミトコンドリア特異的オートファジー:マイトファジーは、ダメージを受けたり、余剰に生じたりしたミトコンドリアを選択的に分解するシステムです。これまでに研究グループは、出芽酵母を用いたゲノムワイドな遺伝子スクリーニングの結果から、GET経路の構成因子を欠損した酵母株でマイトファジーの効率が減少することを見出していました。しかし、Get因子がどのようにミトコンドリアの分解に寄与するかは未解明のままでした。
今回の研究により、Get因子がマイトファジー駆動に必須のミトコンドリア外膜タンパク質Atg32の活性化に必要であることがわかりました。その分子的背景として、Atg32の活性抑制に関わるPpg1-Far複合体がGET経路によって小胞体膜へ繋ぎ止められており、Get因子が存在しないとPpg1-Far複合体はミトコンドリア膜へ局在化し、そこでAtg32の活性化を過剰に抑えてしまうことを発見しました(図2)。
また、ミトコンドリア外膜に配送された「よそ者」タンパク質の除去に関わるMsp1が、Ppg1-Far複合体をミトコンドリア外膜から取り除くことで、マイトファジーの進行に働いている可能性も見出しました(図2)。
以上の知見から、小胞体とミトコンドリアが連携し、マイトファジー活性調節因子の局在を適切に制御することで、ミトコンドリアが過不足なく分解されていると考えられます。マイトファジーは生物進化の過程で高度に保存された基本的な機構であり、本研究を通した出芽酵母での研究成果は、哺乳類マイトファジーの分子理解にも有用な手がかりとなることが期待されます。
図1. 酵母細胞内における膜タンパク質挿入装置と排出装置
GET経路の構成因子のうち、Get3は小胞体膜上のGet1/2複合体へ膜タンパク質を運び込み、Get1/2複合体は膜タンパク質の膜への組み込みを行う。Msp1は、ミトコンドリア外膜に配送された「よそ者」タンパク質の除去に関わる。
図2. マイトファジーにおけるGet因子とMsp1の役割
野生型細胞では、GET経路はPpg1-Far複合体を小胞体膜に繋留している。Get因子破壊株では、Ppg1-Far複合体はミトコンドリア膜へと過剰に局在化し、マイトファジー駆動因子Atg32の活性を抑制する。Msp1は、Ppg1-Far複合体をミトコンドリア膜から除去し、マイトファジーの進行に働いている可能性がある。
研究の背景
ミトコンドリアは外膜と内膜の二重の膜からなる細胞内小器官(オルガネラ)で、主に細胞活動に必要なATPを供給する細胞の「発電所」です。そのため細胞は、エネルギー需要の変動に応じてミトコンドリアの量を増減させる必要があります。またミトコンドリアは、酸化ストレスの過剰な蓄積によりダメージを受け、細胞にとって有害となりうるため、品質の低下したミトコンドリアは迅速かつ適切に分解される必要があります。
ミトコンドリア特異的オートファジー「マイトファジー」は、ミトコンドリアを分解することで、ミトコンドリアの品質や量を調節する機構の一つです。出芽酵母においてマイトファジーが誘導されるとき、他の細胞質成分と区別してミトコンドリアのみを分解の標的とすべく、その目印として働くタンパク質Atg32がミトコンドリア外膜に蓄積します。ミトコンドリア上に集積したAtg32は、Atg8やAtg11などのオートファジー関連タンパク質と結合し、マイトファジーを開始する複合体を形成します。
本研究グループでは、出芽酵母マイトファジーの分子機構の解明を目指してゲノムワイドな遺伝子スクリーニングを行い、マイトファジー関連遺伝子を複数同定してきました。その過程で見出されたのが、膜タンパク質を小胞体膜へ挿入する機能を持つGET経路の構成因子です。
研究の内容
本研究グループは、Get因子を欠損した酵母株において、マイトファジー誘導条件におけるAtg32のリン酸化レベルが減少していることを突き止めました。
Atg32のリン酸化は、Atg11との結合を安定化することが知られており、マイトファジーを駆動させるのに重要です。実際、Get欠損株では、Atg32-Atg11相互作用が不安定化していることがわかりました。
なお、Atg32の脱リン酸化を担い、Atg32-Atg11相互作用の抑制に寄与するタンパク質として、Ppg1が知られています。興味深いことに、Ppg1と結合する小胞体膜タンパク質Farが、Get欠損株ではミトコンドリアへ過剰に標的化する様子が観察されました。この際、Farタンパク質を人工的に小胞体に局在させると、Atg32-Atg11相互作用、マイトファジーが有意に回復することも見出しました(図3)。
これらの結果は、GET経路がPpg1-Far複合体を小胞体膜へ繋留しミトコンドリアへの局在を防ぐことで、Atg32のリン酸化とマイトファジーの駆動に寄与することを示唆しています。
Get欠損株では、種々の小胞体膜タンパク質がミトコンドリア膜へと配送されることが知られています。
一方、ミトコンドリア外膜タンパク質Msp1は、ミトコンドリアに配送された「よそ者」タンパク質を除去する機能を持っています。そこで、GetとMsp1を二重欠損させてみたところ、マイトファジーの効率が相乗的に減少することがわかりました。この際、Farタンパク質を人工的に小胞体に局在させると、マイトファジーが野生型と同程度まで回復しました(図4)。
これらの結果から、Msp1がミトコンドリア外膜上のPpg1-Far複合体を取り除くことで、マイトファジーの進行に働いている可能性が考えられます。
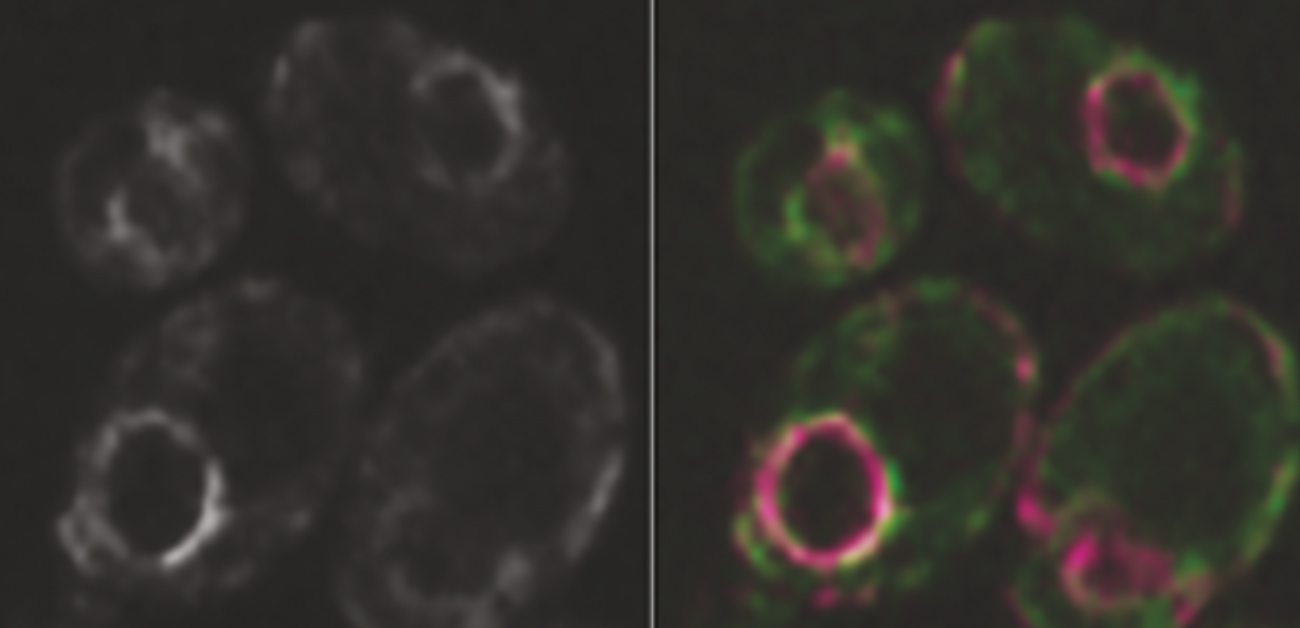
図3. Get因子はPpg1-Far複合体の小胞体局在に機能することで、マイトファジーに寄与する
小胞体とFarタンパク質の共局在を観察した蛍光顕微鏡写真。Get因子破壊株において、Ppg1-Far複合体はミトコンドリア膜へと局在化するが(矢頭)、Farを人工的に小胞体に繋留させることで小胞体局在を回復させると、Atg32-Atg11相互作用、マイトファジーの効率が野生型程度まで回復する。スケールバー、2 µm。
図4. Get因子とMsp1の二重欠損でマイトファジーは低下する
矢印で示すバンドはマイトファジーの効率を示す。Get因子とMsp1の二重破壊株で見られるマイトファジーの低下は、Farタンパク質を人工的に小胞体に繋留させることで見られなくなる。
本研究成果が社会に与える影響(本研究成果の意義)
マイトファジーの駆動因子の活性化機構に対し、小胞体などの異種オルガネラに存在する因子がどのように関わるかはほとんど不明でした。そうした中、本研究では、小胞体の膜タンパク質挿入装置がマイトファジー駆動因子の活性化機構に寄与しているという証拠を得、さらにミトコンドリアの膜タンパク質排出装置と絡めて、マイトファジー制御の一端を明らかにすることができました。こうした視点からマイトファジーの仕組みを解明することは、オルガネラ間連携の新たな役割の理解にもつながり、細胞生物学の分野に広く影響を与えるものと期待しています。
特記事項
本研究成果は、2023年1月25日(水)に米国科学誌「Life Science Alliance」(オンライン)に掲載されました。
タイトル:“The GET pathway serves to activate Atg32-mediated mitophagy by ER targeting of the Ppg1-Far complex”
著者名: Mashun Onishi, Mitsutaka Kubota, Lan Duan, Yuan Tian, Koji Okamoto
DOI:https://doi.org/10.26508/lsa.202201640
参考URL
大阪大学大学院生命機能研究科ミトコンドリア動態学研究室
https://www.fbs.osaka-u.ac.jp/ja/research_group/detail/8
SDGsの目標


用語説明
- ミトコンドリア
ミトコンドリアは、ほとんどすべての真核生物の細胞中に存在する細胞内小器官。脂質二重膜構造を持ち、核とは別途に独自のミトコンドリアDNAを含み、分裂・融合を繰り返し、環境に合わせて動的に変化する小器官である。私たちが生きていくために必要なエネルギー通貨(ATP)を産生する場(発電所)であり、細胞内カルシウムイオン濃度の調整や脂質の合成、免疫反応においても重要な働きを持つことが分かっている。
- マイトファジー
不良または余剰なミトコンドリアを、オートファジーの仕組みを利用して選択的に分解するシステム。マウスや線虫、ショウジョウバエなどを用いた研究から、胚発生や細胞分化時の運命決定、炎症応答、アポトーシス、老化などの多様な現象に関わることが示唆されている。
- 小胞体
細胞質を横断するようにして核膜までつながる、袋上の膜構造によって構成される細胞内小器官。脂質やステロイドの合成、タンパク質の折りたたみや成熟化、カルシウム貯蔵などを担う。
- GET経路
C末端に膜貫通ドメインを持つテイルアンカー型タンパク質を小胞体へ組み込む経路。
- Ppg1
Atg32の不活性化を担う脱リン酸化酵素。主に小胞体に存在するタンパク質Farと結合する。
- Far複合体
Far3-7-8-9-10-11のタンパク質6種から構成される複合体。このうち、Far9, Far10は小胞体膜に組み込まれるテイルアンカー型タンパク質。
- Msp1
ミトコンドリア外膜に存在するAAA-ATP加水分解酵素(ATPases associated with diverse cellular activities)。ミトコンドリア外膜に配送された「よそ者」テイルアンカー型タンパク質を、ATP加水分解に伴い得られるエネルギーを利用して除去する。
- オートファジー
細胞内成分の一部を非選択的あるいは選択的に分解するシステム。二重膜で構成される袋状の膜構造(オートファゴソーム)にタンパク質やオルガネラなどの細胞質成分が閉じ込められ、分解酵素に富んだリソソーム(酵母では液胞)へと輸送され、中身が分解される。