

受精に関わる精子融合因子IZUMO1と卵子受容体JUNOの認識機構を解明
本研究成果のポイント
・哺乳類の受精において精子と卵子の認識に直接関与するタンパク質のIZUMO1とJUNOの立体構造を明らかにしました。
・不明だった精子表面のIZUMO1と卵子表面のJUNOの結合様式が分かりました。
・精子と卵子の認識に直接関わるタンパク質同士の相互作用はこれまで明らかになっていませんでしたが、今回IZUMO1とJUNOの複合体の認識機構が分かったことで、両者の結合を阻害する新たな避妊薬などの開発につながることが期待されます。
リリース概要
東京大学大学院薬学系研究科の大戸梅治准教授、石田英子特任研究員、清水敏之教授、福島県立医科大学医学部附属生体情報伝達研究所の井上直和准教授、大阪大学大学院工学研究科の内山進准教授らの研究グループは、受精において精子と卵子の認識に直接関わるIZUMO1 とJUNO の詳細な立体構造を明らかにしました。
哺乳類の受精では、まず精子が卵子に接着し、その後両者の膜融合が起こることでそれぞれの遺伝情報が組み合わされ遺伝的に新たな個体が作られます。今回立体構造を明らかにしたIZUMO1とJUNOは、それぞれ精子表面と卵子表面に存在するタンパク質で、両者が複合体を形成することで精子は卵子表面に接着します。いずれのタンパク質も受精において必須であることが分かっていましたが、どのような相互作用で結合しているのかは不明でした。
本研究グループは、IZUMO1単体、JUNO単体、IZUMO1-JUNO複合体の3種の立体構造を明らかにしました。その結果、IZUMO1は細長い構造、JUNOは球状の構造をしていることが分かりました。JUNOは葉酸 受容体に非常によく似た構造をしていましたが、葉酸とは結合しないことが知られています。今回の構造から、JUNOは疎水性残基に囲まれたポケットを持っているものの、葉酸認識に重要なアミノ酸残基が一部異なり、またアミノ酸残基の側鎖がポケットを狭める形で配向しているために葉酸と結合できなくなっていることが分かりました。また、IZUMO1-JUNO複合体はIZUMO1とJUNOが1対1の比率で結合しており、IZUMO1の中央部とJUNOの疎水性ポケットの裏側が相互作用することで結合していました。
これらの構造情報をもとに、IZUMO1とJUNOの結合を阻害するような新しい作用機構の避妊薬の開発につながることが期待されます。
研究内容
哺乳類における受精では、精子と卵子が相互に認識し両者の細胞膜が融合することで両者の遺伝情報が組み合わされた新たな個体が作り出されます。受精の過程において、結合や膜融合に関わる因子は特に重要視され、長い間研究の対象として注目されてきました。IZUMO1は精子表面に存在するI型膜タンパク質、JUNOは卵子表面に存在するGPI(グリコシルフォスファチジルイノシトール)アンカー型のタンパク質 で、互いに結合することが分かっています。いずれのタンパク質も欠損させることで受精が起こらなくなることから、精子と卵子の認識において必須の役割を果たしていると考えられます。このため、IZUMO1とJUNOの認識機構を明らかにすることは、基本的な生命現象の解明だけでなく、避妊薬の開発などへの発展も見込まれます。
本研究グループは、IZUMO1とJUNOの相互作用様式をX線結晶解析によって明らかにするために、それぞれの細胞外領域を大量に調製することに成功しました。得られたタンパク質を結晶化し、IZUMO1単体、JUNO単体、IZUMO1-JUNO複合体の3種類の結晶構造を明らかにしました。結晶からの回折データの取得には、高エネルギー加速器研究機構フォトンファクトリーおよび大型放射光施設SPring-8の強力なX線を使用して、2.0~3.2Å(オングストローム、1Åは10ナノメートル)の分解能で構造を決定しました。
構造解析の結果、IZUMO1は細長いロッド状の構造 (図1a) 、JUNOは球状の構造をしていました (図1b) 。また、IZUMO1とJUNOは1対1の比率で複合体を形成しており (図2a) 、基本的にそれぞれ単体の場合と同じ構造でした。IZUMO1は、N末端側の4本のαへリックスの束からなるIZUMOドメイン、中央部分のβヘアピン構造、C末端側のイムノグロブリン様ドメインから構成されていました (図1a) 。中央のβヘアピン構造が主にJUNOとの結合に関与していて (図2a,b) 、この部分はN末端側IZUMOドメインとC末端側イムノグロブリン様ドメインとジスルフィド結合 でつながれることにより安定化されていました (図1a) 。JUNOは葉酸受容体(FRs)と非常によく類似しており、FRsにおける葉酸が結合する部位に相当する疎水性ポケットを持っていました (図1b) 。JUNOは葉酸と結合しないことが知られています。これは葉酸の認識に関与するいくつかのアミノ酸残基がJUNOでは保存されていないことに加えて、ポケット内部の側鎖がポケットを狭める形で配向しているためであることが構造から分かりました (図1b) 。またIZUMO1-JUNO複合体は、IZUMO1の中央のβヘアピン構造とJUNOの疎水性ポケットの裏側で結合しており、互いに相補的な形状で相互作用していました (図2b) 。特に互いのトリプトファン残基が相互作用界面に存在し (図2b) 、これらのトリプトファン残基は他の生物種でも保存されており、基本的な結合様式は共通していると考えられます。実際に、結合に重要だと考えられるIZUMO1の残基に変異を入れた変異体IZUMO1を発現させた細胞は卵母細胞への接着が著しく減少したことから (図2c) 、受精においてIZUMO1-JUNO複合体形成が重要であることが示されました。
本研究により、受精の最初のイベントである精子と卵子の結合がどのような相互作用によって起こっているのかが明らかとなりました。IZUMO1とJUNOの結合を阻害するような新たな非ホルモン性の避妊薬の開発も期待されます。一方で、今回発表された構造は結合の初期段階であり、膜融合に至る過程にはIZUMO1-JUNO複合体の構造変化や、未知の分子が関与している可能性があります。受精のより詳細なメカニズムの解明には、さらなる研究が必要とされます。
本研究は、戦略的創造研究推進事業(CREST)、科学研究費補助金、公益財団法人武田科学振興財団助成金、公益財団法人持田記念医学薬学振興財団助成金、公益財団法人第一三共生命科学研究振興財団助成金などの外部資金支援を受けて行われたものです。
特記事項
発表雑誌 雑誌名:Nature
論文タイトル:Structure of IZUMO1-JUNO reveals sperm-oocyte recognition during mammalian fertilization
著者:大戸梅治 *† 、石田英子 * 、クラユヒナエレナ、内山進、井上直和、清水敏之 † (*共同筆頭著者、†責任著者)
DOI番号:10.1038/nature18596
参考図
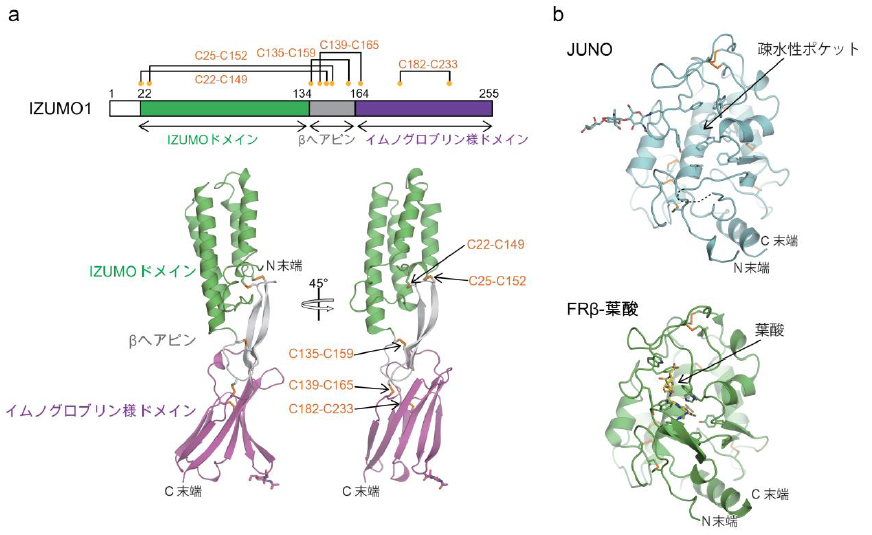
図1 IZUMO1とJUNOの構造
(a)IZUMO1のドメイン構成(上)と構造(下)
(b)JUNO(上)とFRβに葉酸が結合した構造(上、PDB ID:4KMZ)
JUNOを水色、FRβを緑色、葉酸を黄色で示した。
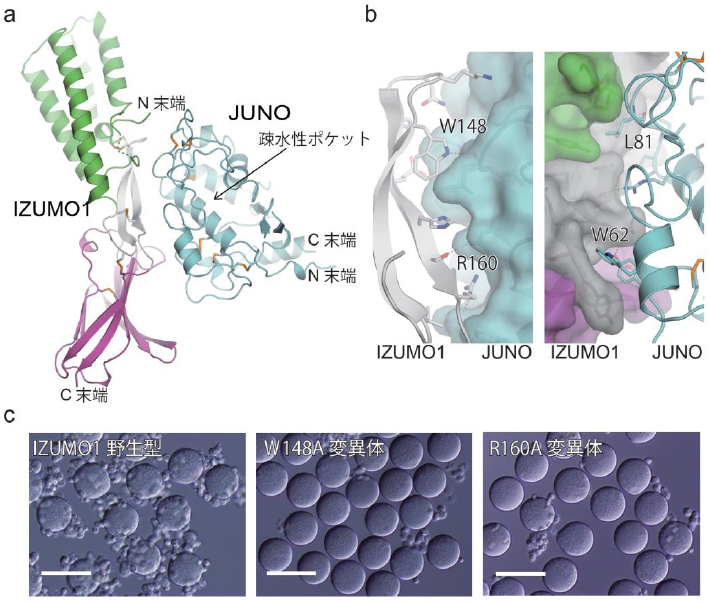
図2 IZUMO1-JUNO複合体の構造
(a)IZUMO1-JUNO複合体の全体図
(b)IZUMO1-JUNO複合体の相互作用界面の拡大図。左図ではJUNO側を表面図(シアン)で、右図ではIZUMO1側を表面図(緑、グレー、紫)で示している。
(c)IZUMO1を発現させた細胞(COS7細胞)と卵母細胞の結合アッセイ。IZUMO1野生型を発現させた細胞は卵母細胞(JUNOを発現している)に接着するのに対して(左)、変異体IZUMO1を発現させた細胞(中央、右)では接着が著しく減少した。白色のバーは100μmを示す。
参考URL
大阪大学大学院 工学研究科 細胞動態学領域 福井研究室HP
http://www.bio.eng.osaka-u.ac.jp/cl/top.php
用語説明
- IZUMO1
縁結びで有名な出雲大社に因んで命名された精子と卵子の融合因子(精子側)。
- JUNO
ローマ神話において結婚と出産を司る女神に因んで命名された精子と卵子の融合因子(卵子側)。
- 葉酸
ビタミンB 9 とも呼ばれる水溶性ビタミン。葉酸からつくられるテトラヒドロ葉酸は、DNAの生合成などに関わる種々の酵素反応の基質として働く。
- GPI
GPI(グリコシルフォスファチジルイノシトール)アンカー型タンパク質:
糖脂質であるGPIがタンパク質のC末端にアミド結合し、細胞膜に繋ぎ止められているタンパク質で、細胞膜表面に局在する。
- ジスルフィド結合
2つのチオールが共有結合したもの。SS結合とも呼ばれる。タンパク質中ではチオール基を側鎖に持つシステイン残基間で形成される。