
植物の気孔と道管の数を調節するペプチド分子を発見!
最適な水輸送機能を形作る仕組みの解明へ
研究成果のポイント
概要
大阪大学大学院理学研究科のチェンピンピン助教、柿本辰男教授らの研究グループは、清華大学(中国)のチャイジジエ教授らの研究グループ、熊本大学の澤進一郎教授らのグループとの共同研究により、モデル植物シロイヌナズナを用いてペプチド性細胞間シグナル分子CLE9/10(以降、CLE9/10)の機能を解明しました。
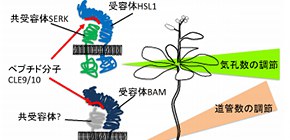
植物の道管と気孔は共に水の通り道です。今回、CLE9/10が受容体HSL1・共受容体SERK複合体に結合して気孔数を制御すると共に、受容体BAMに結合して道管数を調節していることを明らかにしました (図1) 。CLE9/10は別々の受容体を介してこれらの数を制御することにより、最適な水輸送機能を持つ植物の形を作っていると考えられます。
本研究成果は、2018年12月5日(英国時間)に英国科学誌「Nature Plants」にオンライン公開されました。
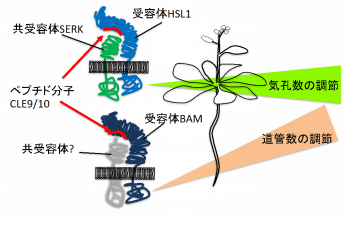
図1 一つのペプチド性細胞間シグナル分子CLE9/10が、活性化する受容体によって違った発生過程を制御する。
研究の背景
植物は、根から葉までつながる道管と、葉の表皮にある気孔という小さな穴を使って水分を調節しており、発生過程における道管と気孔の数の制御は重要な役割を担っています。根端には将来道管になる前駆細胞があり、各前駆細胞が根の長軸と直角な方向に細胞分裂することによって道管の細胞列を作り出し、最終的には細胞が中空になると共に、細胞列を作る細胞間が貫通することで道管が作られます。道管前駆細胞の数は、道管前駆細胞の縦方向の細胞分裂によって増やされますが、その制御機構は良くわかっていませんでした。
葉の表皮には幹細胞であるメリステモイド母細胞(MMC) が存在します。MMCは全ての気孔孔辺細胞と多くの一般の表皮細胞の源となっています。MMCの幹細胞としてのアイデンティティーを決めている転写調節因子はSPCHです。これまでに、ペプチド性細胞間シグナル分子EPF2がMMC数の制御に働いていることは柿本教授らのグループにより明らかにされていましたが、MMC数はさらに別の細胞間シグナル分子にも制御されているだろうと考えられていました。
研究の成果
植物の発生は植物ホルモンと多くのペプチド性細胞間シグナル分子によって制御されています。ペプチド性細胞間シグナル分子のうち、CLEグループのペプチド(CLEペプチド)はシロイヌナズナには32種類存在します。発表者らは、CLE9/10(CLE9遺伝子とCLE10遺伝子によって同一のCLE9/10がコードされる)と呼ばれる13アミノ酸からなるペプチドが、気孔と、気孔以外の表皮細胞の数を制御していることを見出しました。CLE9/10がどのような受容体にとって認識されているのかを知るために多くの候補遺伝子の破壊株のCLE9/10への応答性を調べ、細胞膜に存在する受容体キナーゼHSL1がその受容体であることを見出しました。HSL1がCLE9/10を認識すると、SERKと呼ばれる共受容体を呼び込んで細胞内に情報を伝え、その結果、葉の幹細胞であるMMCのアイデンティティーを支配する転写調節因子SPCHを消失させ、気孔幹細胞の数を抑制することが明らかになりました。
CLE9遺伝子は道管前駆細胞でも発現しています。MMCにおいてはCLE9/10はHSL1によって受容されていましたが、道管前駆細胞においてはCLE9/10は別の受容体キナーゼであるBAMにより認識されて道管前駆細胞数を制御していることがわかりました。
本研究成果が社会に与える影響(本研究成果の意義)
植物は多くのペプチド性細胞間シグナル分子候補と受容体キナーゼを持っています。これらは必ずしも1対1対応ではなく、1つのペプチドが違った受容体を介して違った機能を発揮する例を示したものです。ここでは、一つのペプチドが、水の通り道である道管と気孔数を、別の受容体を介して調節していることを示したものです。さらに、受容体BAMには複数のペプチドが作用することを本研究グループや他の研究者が見出しており、多くのリガンドと受容体が複雑な相互作用関係を持っていることがわかります。
研究者のコメント
植物も動物と同様、発生の制御のために、細胞同士はペプチドを含む数多くのシグナル分子を用いてコミュニケーションしています。今後の研究の発展により、その全貌が次第に明らかになってくると考えています。
特記事項
本研究成果は、2018年12月5日(英国時間)に英国科学誌「Nature Plants」にオンライン公開されました。
タイトル:“The CLE9/10 secretory peptide regulates stomatal and vascular development through distinctreceptors”
著者名:Pingping Qian 1* , Wen Song 2,3,4* , Toshiya Yokoo 1 , Ayako Minobe 1 , Guodong Wang 1,5 , Takashi Ishida 6 , Shinichiro Sawa 6 , Jijie Chai 2,3,4# , Tatsuo Kakimoto 1#
*equal contribution #責任著者
1.大阪大学、2.清華大学(中国)、3.Max Plank研究所(ドイツ)、4.ケルン大学(ドイツ)、5.陝西大学(中国)、6.熊本大学
なお、本研究は科学研究費補助金基盤研究(B)((25291060)(柿本))、新学術領域研究(25113006、18H04837(柿本))およびMinistry of Science and Technology of China(2015CB910200(Chai))の支援により行われました。
参考URL
大阪大学 大学院理学研究科 生物科学専攻 植物生長生理研究室
http://www.bio.sci.osaka-u.ac.jp/bio_web/lab_page/cell_physiol/sitepg/Kakimoto_Lab/homu.html
用語説明
- 細胞間シグナル分子
細胞間の情報伝達を担う分子のこと。
- 受容体
情報伝達など関わる分子を受容するタンパク質を一般に受容体という。ここでは、受容体キナーゼ のこと。
- 受容体HSL1
植物は不要になった花びらなどを脱離させますが、脱離を促すペプチド情報を受け取る受容体としてHAESAがあります。HAESAに似た受容体としてHSL1(HAESA-like1の略)が知られていましたが、本研究まではその機能がわかっていませんでした。
- 共受容体SERK
植物の受容体の多くは、情報分子を受け取ると、共受容体と相互作用することで細胞内に情報を伝えます。SERKに分類される共受容体は、植物ホルモンの一種であるブラシノステロイドの受容体や病原体を認識する受容体など、様々な受容体と相互作用して細胞内に情報を伝えます。
- 受容体BAM
シロイヌナズナはBAMに分類される3つの受容体と、これらに良く似た受容体CLV1を持っています。これらの受容体は茎頂に存在する幹細胞群の数を決めるなどの役割が知られていましたが、本研究では道管の数も調節していることが分かりました。
- メリステモイド母細胞(MMC)
葉原基の原表皮細胞の一部が転写因子SPCHの作用でMMC(メリステモイド母細胞)になります。MMCは不等分裂をして三角形のメリステモイドとその姉妹細胞となります。メリステモイドも姉妹細胞もSPCHが十分に働いているとMMCの機能をもち、再び不等分裂をしてメリステモイドとその姉妹細胞となることができます。最終的にはメリステモイドは孔辺細胞母細胞となった後に等分裂をして気孔孔辺細胞となり、メリステモイド以外の細胞は一般的な表皮細胞となります。最終的な気孔と表皮細胞の数は、MMCの数とMMCとして維持される期間に依存します。
- SPCH
SPCHは気孔細胞系譜の幹細胞のアイデンティティーを与える重要な転写調節因子です。植物は細胞外の情報をもとにSPCHの量を調節し、これによって気孔やそれ以外の表皮細胞の数を調節していることが知られています。
- CLEペプチド
植物の分泌性ペプチド。CLAVATA/EMBRYO SURROUNDING REGION の略。最初に報告されたペプチドは茎頂分裂組織の細胞増殖の負の調節因子CLAVATA3です。
- 受容体キナーゼ
細胞外にシグナル分子認識ドメイン、細胞内にリン酸化酵素(キナーゼ)ドメインを持ちます。植物には非常にたくさんの受容体キナーゼが存在し、細胞間のコミュニケーションや、病原体の認識などを行っています。