
染色体の“駆動エンジン”を新しい場所に載せ替える仕組みを解明
柔軟かつ頑強な染色体の性質が明らかに
リリース概要
大阪大学大学院生命機能研究科の石井浩二郎招へい准教授の研究グループは、各細胞に含まれる染色体の駆動エンジンとなる動原体が、染色体上の新しい場所に設置される手順を新たに明らかにしました。動原体は細胞分裂期の染色体の精緻な動きを作り出す重要な高次構造体です。染色体を正確に分配しゲノムを確実に継承していくために、動原体は各染色体に必ず一つずつ必要であり、それはセントロメアと呼ばれる特殊染色体領域に設置されるのが基本です。しかし今回の研究では、セントロメアではない染色体領域にも動原体は安定に設置可能であり、そのためには設置領域から染色体の基礎構成要素であるヒストン蛋白質のある特定型(H2A.Z)が排除されることが条件であることが明らかとなりました。基本以外に例外を認めるこのような染色体の仕組みは、不測の事態にも柔軟に対応できる染色体の頑強さを表すものです。今回の発見は、その頑強な染色体特性の分子レベルでの理解に向けた重要な一歩であるといえます。
なお、本研究成果は11月3日13:00(米国東部時間)に米国科学雑誌「Nature Structural & Molecular Biology(ネイチャー・ストラクチュラル・アンド・モレキュラー・バイオロジー)」オンライン速報版で公開されます。
研究の背景
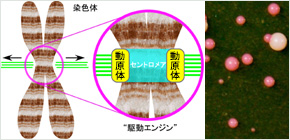
染色体の多くは細胞分裂期には中央部がくびれた「X型」のかたちを呈します。そのくびれ部分には、染色体の駆動エンジンである「動原体」が形成されます( 図1 )。実際には、くびれが染色体の端にある場合や、くびれを全く示さない染色体も知られていますが、どの染色体にも動原体は必ず一つ存在します。なぜなら、染色体は生命の設計図であるゲノムDNA を収納しており、動原体が作り出す染色体の緻密な動きは、ゲノムを娘細胞に忠実に継承するために必要不可欠だからです。
くびれ部分は「セントロメア」という特殊な繰り返しDNA配列で成り立っています( 図1 )。しかし近年の研究により、動原体はセントロメアDNA以外のDNA配列上にも形成されうることが分かってきました。そのような非セントロメアDNA配列上での動原体の形成は、偶発的に生まれたセントロメアを持たない染色体断片を安定に維持させる正の働きがありますが、同時に、複数の駆動エンジンを一つの染色体上に作り出して染色体の適正な動きを損なわせ、ゲノムを不安定化してしまう負の働きもあります。その働きがどのように調整されているのか、これまではよく分かっていませんでした。
石井准教授のグループは、酵母を操作して生きたまま一つの染色体のセントロメア領域を破壊する実験を進めてきました( 図2 )。セントロメア破壊によって大半の酵母は死滅しますが、その中からセントロメア以外の新しいDNA領域に動原体を形成して生き残ってくる酵母細胞が見出されます。今回、第三染色体のセントロメア破壊実験を行ったところ、他の染色体の場合と同様に生き残る酵母は見出されましたが、その細胞は病的で、出現後しばらくすると死んでしまうことが分かりました。そして、細胞が病的なのはその細胞の中の第三染色体上に新たに設置された動原体がまるで“仮”設置のように不安定なことが原因であり、その動原体の不安定さはH2A.Zという特定型のヒストン蛋白質 を含む染色体領域に動原体が仮設置されたために動原体の安定維持に必要な蛋白質の結合が低下した結果生じていることを明らかにしました( 図2 )。ヒストンH2A.Zを含まない領域に設置された動原体は問題なく安定に維持されます。また不安定だった動原体もヒストンH2A.Zを設置領域から除去すると安定に維持されるようになりました( 図3 )。
本研究成果が社会に与える影響(本研究成果の意義)
ヒストンH2A.Zは遺伝子発現の高い染色体ゲノム領域により多く蓄積することが知られています。今回の発見は、染色体上の重要な遺伝子発現領域が動原体の形成領域と重複してしまうことを避けるための保証機構を明らかにしたものと言えます( 図2 )。この機構によって染色体駆動エンジンの“無軌道”な設置は避けることができます。一方で、不安定な動原体のみに駆動を頼ることになった染色体は、細胞に病的さを与えますが即死にするわけではなく、そこから正常に生育する適応細胞を生み出す猶予を作り出しています( 図3 )。秩序は保ちつつも硬直しない、染色体機能制御の適度に柔軟な側面が如実に感じられる機構です。生物進化さえも下支えすると思われるこの染色体の頑強な特性を、今後分子レベルで解明していくのに大いに役立つ発見と考えます。また、新規DNA領域での動原体形成はこれまでに様々な腫瘍細胞で見つかっています。腫瘍形成とゲノム不安定性の間には深い関連が指摘されており、本研究成果は将来的な制がん治療の技術開発に関わってくる可能性もあるものと考えます。
特記事項
本研究成果は、科学研究費補助金・新学術領域研究「ゲノムアダプテーション」や基盤研究(B)などによる助成の一環として得られました。大阪大学生命科学研究独立アプレンティスプログラムの支援も受けています。
論文掲載情報
Yuki Ogiyama, Yuko Ohno, Yoshino Kubota, and Kojiro Ishii (荻山友貴、大野悠子、久保田佳乃、石井浩二郎) Epigenetically induced paucity of histone H2A.Z stabilizes fission yeast ectopic centromeres (エピジェネティックに誘導されたヒストンH2A.Zの欠如が分裂酵母の異所的セントロメアを安定化する)
参考図

図1 染色体と動原体の模式図
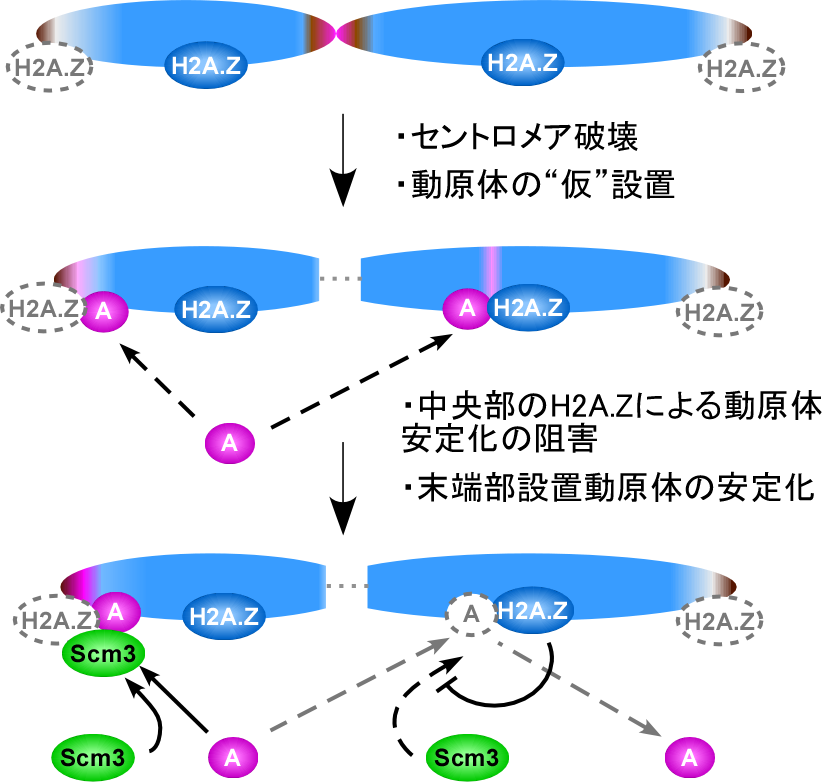
図2 セントロメア破壊染色体の新規DNA領域に動原体が安定に設置される手順
(上段)染色体のセントロメア部分と末端部分はヒストンH2A.Z(青)が少ない。 (中段)セントロメアを破壊すると、動原体(マゼンタ楕円“A”)は一過的には末端部分にも中央部分にも“仮”設置される。 (下段)しかし、H2A.Zの存在する中央部分では動原体の安定維持に必要な蛋白質(緑楕円“Scm3”)が安定に結合できないために仮設置された動原体はそのうちに消滅するが、H2A.Zのない末端部分ではScm3が安定に結合し、動原体Aの安定な設置が完了する。第三染色体では末端部分の動原体形成領域にも初めからH2A.Zが存在しているため、中段の“仮”設置状態の動原体しか見出されない。

図3 不安定な動原体をもつ酵母(赤)から安定な動原体をもつ酵母(白)が出現する
白い酵母コロニーは赤い酵母コロニーから自発的に生まれてくる。白コロニー中に含まれる酵母細胞では第三染色体がさらに変化を起こしており、その動原体の設置領域からはヒストンH2A.Zがなくなっていた。
参考URL
大阪大学大学院生命機能研究科 染色体機能制御研究室
http://www.fbs.osaka-u.ac.jp/labs/ishii/
用語説明
- ゲノムDNA
生物が持つ全デオキシリボ核酸(DNA)の総体。遺伝子のもつ情報はゲノムDNA中に暗号配列として全て書き込まれている。各遺伝子の配列情報を元に細胞内で作り上げられる蛋白質などの実際の働きが、さらに高度に組み合わさって複雑な生命プログラムは遂行されるため、ゲノムDNAはその設計図と表現される。
- ヒストン蛋白質
染色体がゲノムDNAを折り畳んで収納するのにあたり、最も基礎的な折り畳みを担う蛋白質。H3, H4, H2A, H2Bの四種類のヒストン蛋白質が二分子ずつ結合した構造が芯となって、ゲノムDNAをその周りに約二回転巻き取る。ヒストン蛋白質によるDNAの巻き取りは基本的にゲノムDNA分子の端から端まで均一に起きている。ヒストンH2A.ZはヒストンH2Aの異型蛋白質。